Смотреть другой контент
Рубрики Все →
Поиск статей
Последние статьи
- Тайна пещеры "Путь к коммунизму...
- Всероссийская Научная Конференция «Пути Эволюционной Географии», Посвященная Памяти Профессора Андрея Алексеевича Величко. Ответ рецензентам
- Ответ на рецензии
- КЛИМАТИЧЕСКИЕ КАТАСТРОФЫ В ИСТОРИИ ЧЕЛОВЕЧЕСТВА. ВОЗРАСТ АРХЕОЛОГИЧЕСКИХ СООРУЖЕНИЙ ДРЕВНЕГО ЕГИПТА ПО ПАЛЕОГЕОГРАФИЧЕСКИМ ДАННЫМ
Последние коментарии
-
 Тайна пещеры "Путь к ко...
Тайна пещеры "Путь к ко...В.Юрковец - Дек 19 2023 03:12
-
Филогенетическое древо R1a1...
В.Юрковец - Янв 04 2017 01:04
-
Климатические корреляции в...
В.Юрковец - Янв 29 2015 02:14
-
О статье Божидара Митровича...
В.Юрковец - Ноя 14 2014 19:44
-
 О статье Божидара Митровича...
О статье Божидара Митровича...Владимир - Сен 27 2014 01:00
-
О статье Божидара Митровича...
Владимир - Сен 27 2014 00:03

СЕ - ЧЕЛОВЕК
Мар 24 2013 10:05 |
В.Юрковец
в Статьи А.Клёсова
 CE – ЧЕЛОВЕК
CE – ЧЕЛОВЕКА.А. Клёсов
ЧАСТЬ ПЕРВАЯ – О МУЖЧИНАХ
Разрешите представиться. Я - 13-24-16-11-11-15-12-12-10-13-11-30. Это по отцовской линии. По материнской - 16519С, 263G, 309.1C, 315. 1C, 477C.
Это - то, что я на самом деле имею. Это мой настоящий паспорт. Моя идентификация. Мой гаплотип. Все остальное в жизни я мог выбрать - по своему желанию или по обстоятельствам. Но не наследственный код, записанный в моей ДНК.
Эти два ряда цифр уходят во тьму веков и тысячелетий. Первый набор цифр я получил от своего отца, он - от своего отца, тот - от своего, моего прадеда. И так далее, на глубину примерно 160 тысяч лет. Туда, к моему предку 160 тысяч лет назад, сходятся все прямые генеалогические нити от всех живущих сегодня мужчин. Он - прямой праотец всех мужчин на сегодняшней Земле.
Правда, в последнее время картина с первопредком усложнилась. Оказалось, вопреки тому, что нам твердили последние двадцать лет, наши предки вышли не из Африки. Кроме того, в Африке были обнаружены еще ДНК-линии, которые уходят гораздо глубже 160 тысяч лет назад, возможно, что и на глубину 300 тысяч лет назад или больше, и не имеют к нам, европейцам, вообще почти никакого отношения. Вот они, по-видимому, и есть настоящие древнейшие африканцы, но мы от них не произошли.
Тем не менее, первопредка остальной части человечества условно называют "хромосомным Адамом". Конечно, он не был первым человеком, первым мужчиной на Земле. Просто остальные, кроме его прямых предков, в том числе старше его на тысячелетия, на десятки и сотни тысяч лет, не выжили, не оставили мужского потомства, или потомство оборвалось на протяжении этих тысячелетий. Войны, сражения, убийства, болезни, рождение только дочерей, ранняя смерть сыновей - все это терминирует прямую мужскую генеалогическую линию. Тысячи таких терминирований происходят сегодня, каждый день. Но тысячи каждый день получают продолжение, с рождением каждого мальчика.
Первая цепочка цифр передается только мужчинам. Они, эти цифры, передают в привычной для нас арифметической системе определенные последовательности нуклеотидов в нашей мужской хромосоме Y, которой нет у женщин. Эти последовательности очень консервативные, и не мутируют многие поколения. Они записаны в так называемых "никчемных" последовательностях хромосом, которые ничего не кодируют. Но они, эти последовательности, с поразительной точностью копируются от отца к сыну, из поколения в поколение.
У женщин - своя система. Второй ряд цифр, приведенный в самом верхнем абзаце, передает расположение нуклеотидов в короткой цепочке митохондриальной ДНК. Какой такой митохондриальной ДНК - об этом речь позже. Главное - что эти последовательности передаются тоже с поразительной точностью от матери - дочери. Дочь получила их от своей матери, та - от своей. И так далее, на глубину примерно 180 тысяч лет. Туда, к "митохондриальной Еве" сходятся все прямые генеалогические нити от всех живущих сегодня людей. Она - прямая прамама всех людей на сегодняшней Земле. Да, и женщин, и мужчин. Потому что та цепочка ДНК, передающаяся от матери дочери, передается в том же виде и сыну.
Так что наша Ева никогда не встречала нашего Адама. А почему митохондриальная Ева старше хромосомного Адама - на то есть причины. Женщины в целом дольше живут. Их меньше выбивали на войнах. И, самое главное - что женская генеалогическая информация хотя и терминируется на мальчиках, но девочкам-то передается, а те передают ее своим детям, как девочкам, так и мальчикам. Так что сыновья эту информацию хоть и не передают, но сохраняют.
Все люди на Земле - генетические и генеалогические родственники. Все (или почти все) произошли от одного предка по мужской линии, и от одной - по женской. Остальные, как уже было сказано, не выжили. Все мужчины на Земле, все 100%, имеют вот такой набор маркеров в той самой «никчемной» части ДНК:
DYS 19, 388, 390, 391, 392, 393
Это – минимальный набор, который обычно рассматривают в ДНК-генеалогии. Но на самом деле маркеров в ДНК-генеалогии гораздо больше, несколько сотен. Все они имеют присвоенные им номера, которые особенного смысла не имеют. Просто номера по принятой классификации. Эти «маркеры» еще называют «локусы», и с ними связаны такие понятия, как «микросателлиты», или STRs, short tandem repeats. Почему repeats, то есть повторы, они же аллели – об этом чуть ниже.
Этот набор еще называют «научным», потому что именно его на заре ДНК-генеалогии, всего несколько лет назад, использовали в научных публикациях. В ДНК-генеалогии чаще используют 12-маркерный набор, например, тот, который приведен в самом начале этого рассказа. Для более детальных описаний – 25-маркерный, 37–маркерный, 67-маркерный, или даже 111-маркерный, который сейчас становится наиболее популярным. В принципе, каждому мужчине можно выдать «паспорт», состоящий из сотен таких цифр, но это пока не имеет практического значения.
Эти наборы сохранились в наших ДНК от самых первых мужчин, которые смогли передать нам всем свою генеалогическую память, и потомство которых выжило 160 тысяч лет назад, и пронесло эти маркеры до настоящего времени.
DYS расшифровывается как "ДНК Y Сегмент", по-английски DNA Y Segment. Y - это та самая мужская половая хромосома, о которой речь шла выше. Мою ДНК анализировали по 111 маркерам, первые двенадцать из которых приведены в начале статьи. Я мог бы привести и все 111 цифр, но всему свое время.
Хотя, впрочем, пожалуйста:
13 24 16 11 11 15 12 12 10 13 11 30 16 9 10 11 11 24 14 20 34 15 15 16 16 11 11 19 23 15 16 17 21 36 41 12 11 11 9 17 17 8 11 10 8 10 10 12 22 22 15 10 12 12 13 8 15 23 21 12 13 11 13 11 11 12 13 31 15 9 15 12 25 27 19 12 12 12 12 10 9 12 11 10 11 12 30 12 14 25 13 9 10 18 15 20 12 24 15 12 15 24 12 23 19 11 15 17 9 11 11
А что такое "анализировали"? - спросит любознательный читатель.
А вот что. Сама хромосома Y - относительно невелика по сравнению с другими хромосомами человека. Все 46 хромосом, 23 пары, в совокупности состоят из трех миллиардов нуклеотидов, и в их составе примерно 30 тысяч генов. Стало быть, в среднем по 65 миллионов нуклеотидов и по 652 гена на хромосому. Хромосома Y состоит из 50 миллионов нуклеотидов, и в ней - всего 40 генов. Остальная часть - некодирующая, "никчемная". Оказалось, в этой "никчемной" части имеются некие загадочные повторы нуклеотидов. Например, шестнадцать раз повторяется квадруплет ТАГА (тимин - аденин - гуанин - аденин). Вот так:
ТАГАТАГАТАГАТАГАТАГАТАГАТАГАТАГАТАГАТАГАТАГАТАГАТАГАТАГАТАГАТАГА
Или - одиннадцать раз триплет ТАТ (тимин - аденин - тимин). Ну, суть ясна. Оказалось, что эти повторы могут служить генеалогическими маркерами. Потому что эти повторы одинаковы у меня и моего отца. И моего дедушки. И дедушки его дедушки. И так далее, в ту самую глубину веков. А у другого человека, не моего прямого родственника, число этих повторов уже другое, хотя сам маркер - тот же.
На самом деле, если пройти в глубь веков, число повторов будет немного различаться, и именно это делает возможной ДНК-генеалогию. Потому что если бы у всех было одинаково, то и у любого другого человека было бы то же самое. И тогда не было бы ДНК-генеалогии. Но в жизни абсолютного постоянства нет. Об этом немного позже.
Число повторов в генетике называют аллелями, но не будем насыщать этот рассказ научными терминами. Пусть будут повторы.
Квадруплетный маркер ТАГА (в повторах) носит номер DYS19. Все мужчины на Земле имеют этот маркер, но в повторах от 11 до 19 раз подряд (их называют "тандемные повторы"). У меня этот маркер повторяется 16 раз подряд (третий по счету в моем гаплотипе выше). А, скажем, у Томаса Джефферсона, третьего президента США - 15 раз. То есть по этому одному маркеру мы с Джефферсоном удалены генеалогически на один шаг. По другому маркеру, DYS392, повторяющемуся триплету ТАТ, у меня 11 повторов подряд (11-й по счету в моем гаплотипе). У Джефферсона, раз уж мы взяли его за сравнение - 15 раз. То есть дистанция выросла до пяти шагов только по двум маркерам. Из этого уже можно сразу сказать, что он не мой близкий родственник. В смысле, что наш с Джефферсоном общий предок жил десятки тысяч лет назад, и ближе по времени общих предков у меня с ним уже не было. Потому что для «научных» наборов из шести маркеров дистанция в один шаг – это уже в среднем три тысячи лет до общего предка. А тут на двух маркеров пять шагов – десятки тысяч лет разницы.
Так вот, генеалогический анализ хромосомы Y - это и есть определение числа повторов в каждом маркере. Результатом является гаплотип конкретного человека, например, в том виде, который записан в самом начале этого рассказа, и расширен чуть выше. Раньше анализ проводили по по 12 или 25 маркерам, но уже не проводят, сместились к более протяженным, 37 маркерам (сейчас это стоит в американской тестирующей компании 169 долларов, хотя недавно было 260 долларов) или 67 маркерам (сейчас – 268 долларов, снизилось с 350 долларов), но для того, чтобы платить такие ощутимые деньги, нужно понимать, что эти анализы могут дать и зачем они нужны.
Самая известная компания, которая делает эти тесты – FamilyTree DNA, которая находится в Техасе, США. http://www.familytreedna.com/
Итак, подводим промежуточный итог. "Мужской" генеалогический анализ, или определение гаплотипа, можно проводить по определенному набору маркеров. Чем набор больше, тем точнее, но и дороже. Результат даст вам возможность найти своих родственников, имеющих одного общего предка, и оценить, насколько этот наш общий предок удален во времени. Масштаб удаления - сотни лет, тысячи, десятки тысяч лет, в зависимости от "генеалогической дистанции".
Но надо понимать, что найти родственников по одному своему гаплотипу – это что искать иголку в стоге сена. Может и повезти, конечно, но лучше использовать иголкоискатель. А точнее – профессиональные методы ДНК-генеалогии, которыми владеют специалисты.
А почему так, спросит читатель? - Ведь даже 12-маркерный гаплотип – это, говорят, почти уникальный. Хорошо, разберемся, что такое здесь «почти». В 12-маркерном гаплотипе, приведенном в самом начале рассказа – а это типичный «славянский» гаплотип жителей России, Украины, Белоруссии, Польши, в также многих литовцев, чехов, словаков, немцев, индусов – так что не такой уж он и «славянский», как выясняется, потому и взят в кавычки, так вот, в нем половина аллелей (это – цифры в нем) почти у всех одинаковы. Это и первая 13, и четвертая 11, и седьмая 12, и восьмая 12, и десятая 13, и одиннадцатая 11. У других этносов (а, точнее, гаплогрупп) – другие, а у половины мужчин-славян России, например, практически одинаковы. И у половины поляков, и у половины украинцев и белорусов, и у одной шестой индусов, то есть на круг только перечисленных – 150 миллионов человек. И у подавляющего большинства этих 150 миллионов вариации идут только по шести маркерам из 12. Если приложить немного математики, то вероятность совпадения по всем 12 маркерам – примерно 6%. То есть у шестерых из 100 – совпадут. Вывод – из 150 миллионов человек у 9 миллионов будут одинаковые 12-маркерные гаплотипы. Вот и ищите родственников.
А если перейти на 67-маркерные гаплотипы, то совпадения будут только у семерых из этих 150 миллионов человек Совсем другое дело. Иначе говоря, случайного совпадения просто не может быть. Если совпало, или в пределах одной-двух мутаций – точно родственник, в пределах 200-400 лет до общего предка. Потому и искать родственников надо на 67-маркерных гаплотипах, а не на 12-маркерных.
А почему число повторов в маркерах разное, и какое это имеет отношение к удаленности общего предка? А то, что чем дальше предок во времени, тем большее количество мутаций произошло в его хромосомах, и, в частности, в тех самых маркерах. Фермент, считывающий число повторов в зоне маркеров, работает исключительно точно. Представляете - пилит фермент вдоль квадруплетного повтора ТАГА, каждая из идентичных четверок повторяется, скажем, шестнадцать раз (DYS19 = 16), в сумме 64 однообразных остатка, и наш фермент ни разу не ошибается, воспроизводя - реплицируя - эту тандемную цепочку. То есть не ошибается пятнадцать поколений, около четырехсот лет. И вдруг бац - ошибся. Напутал. Повторил не 16 раз, как завещали предки, а 15 или 17 раз. Мутация. Shit happens, как говорят американцы.
И родился мальчик с другим числом повторов данного маркера, сам того не подозревая. И, когда подрастет, родятся у него мальчики уже с DYS19 = 15 или DYS19 = 17, смотря по тому, в какую сторону произошла мутация. И если племя этого мальчика, ставшего к тому времени мужем, и оставившего мужское потомство, успешно передвинется в новые края, то вместе с ними передвинется и DYS19 = 17, по пути приобретая новые мутации в среднем одну на каждые несколько поколений. Так образуются гаплогруппы - характерные картины повторов в маркерах ДНК, присущие группам населения, плюс дополнительные метки-мутации.
При этом маркеров много, и мутации в каждом происходят с разной вероятностью во времени. В каждом маркере как бы свои часы, со своим текущим положением стрелок. И вот совокупность этих стрелок по всем маркерам и дает индивидуальность того «паспорта», о котором шла речь в самом начале. Да еще и циферблаты разные – у каждого рода – свой. Эти разные циферблаты и есть дополнительные метки-мутации. То есть у каждого рода – своя гаплогруппа.
Чем более изолирована группа - тем более характерна и четка гаплогруппа. Но даже и при перемешивании групп населения гаплогруппы достаточно легко прослеживаются, поскольку отражают гаплотипы наших далеких предков, живших тысячелетия назад. То есть люди перемешались, а их гаплотипы - нет. Они, повторяю, неизбежно передаются из поколения в поколение.
Так что добавим к нашему "промежуточному итогу" - помимо возможностей найти родственников по прямой нисходящей мужской линии и оценить, примерно когда жил наш общий с найденным родственником предок - еще возможность найти, к какой исторической общности людей, к каким племенам относились наши предки. Современная наука пока не знает, откуда вышли наши самые далекие предки вида Homo sapiens, «человек разумный», под «подозрением» - и Европа, и Ближний Восток, и Азия, некоторые продолжают считать, что Африка – но это самая слабая гипотеза. В любом случае люди стали расходиться от места формирования человека разумного, или формировали его на ходу, в ходе миграций, и третьи двинулись кто в Европу, кто в Сибирь, кто по северной части, перейдя Берингов пролив (в то время - посуху), распространились по Америкам, иные прошли по низам Гималаев в нынешние Китай и Юго-Восточную Азию, и оттуда переправились в Полинезию. Именно анализ гаплотипов показал, что неправ был Тур Хейердал, не из Южной Америки, а из Азии заселилась Полинезия.
И генеалогический анализ нашей Y-хромосомы позволяет ясно видеть отблески тысячелетней истории наших предков. Не предков вообще, а наших, каждого в отдельности прямых предков. Наших прародителей.
История в лицах
К настоящему времени база данных по маркерам Y-хромосомы составляет примерно 500 тысяч данных. Пока – капля в море, меньше одной десятой процента мужского населения мира. Но уже с помощью компьютера можно получить интересные данные. Мои первые 12 маркеров были введены в эту базу данных, и компьютер показал восемнадцать человек с абсолютно точным совпадением по всем 12 маркерам и по повторам в каждом из них. То есть этот мой конкретный гаплотип делят со мной 17 человек. Кто такие? Имена - вовсе не Иванов-Петров-Сидоров. Хотя есть Окулов, Арбузов, Фефелов, Шупенко. С другой стороны, по 12 маркерам наш общий предок всех 18 человек - а что он был, и относительно недавно в историческом масштабе лет, несомненно - мог жить и пять тысяч лет тому назад. То что он был – несомненно, потому что все 18 человек относятся к одному роду – R1a, а у каждого рода был общий предок. Более строгие расчеты показывают, что совпадение двух 12-маркерных гаплотипов помещает общего предка этих двух человек на 625±625 лет назад. С одним человеком из этого списка у меня две мутации на 25-маркерных гаплотипах, то есть общий предок жил 550±390 лет назад. Как такие данные рассчитываются, я здесь пояснять не буду, иначе это усложнит рассказ. В общем, имена тех, у кого со мной полное совпадение по 12 маркерам:
Виктор Окулов (Россия)
Владимир Арбузов (Россия)
Александр Шупенко (Россия)
Фефелов (Россия)
Timothy Meese (Англия)
Wayne Henderson (Англия)
Michael Halfman (Германия)
James Cashner (Эльзас-Лотарингия, Германия)
David Schmidt (Gohrau, Германия)
Cedric Grifin (Англия)
Alvin Benson (Норвегия)
Lukasz Burczynski (Польша)
G. Moga (Индия)
Donald Stonefeld (Англия)
Serge Lariviere (Канада)
Wagner (страна неизвестна)
Mustafa Camlica (Турция)
Понятно, что если бы охват по тестируемым был не 30 тысяч человек (столько тестировались на гаплогруппу R1a), а 30 миллионов, то список был бы в тысячу раз длиннее. Но пока что есть, то есть. Да, еще есть Эрик Римехауг, норвежец, с кем у меня два отклонения на 16 маркерах, и три отклонения – на 21 маркерах. Его самый далекий известный прямой предок Эйкли Хермунд родился в 1515 году в городке Гол в Норвегии, прожил 65 лет.
Ну, хорошо. А как насчет того, чтобы проверить гораздо более жестко - по 25 маркерам? Сузить поиск?
Оказался родственник - один, и то с двумя шагами (мутациями) в сторону. Что для 25 маркеров - просто почти родной. Кстати, это объясняет, что 37 маркеров мне пока в этом отношении пользы не принесли. Как и 67 маркеров. С 25-ю - и то не густо.
Да, к моему родственнику, Михаэлю Хафману. Живет в Германии. Его наиболее удаленный во времени предок, о котором имеются достоверные сведения - Хейнрих Хафман, родился в 1790 году в Германии. Прожил, чертяка, в те времена 70 лет. Статистически, с учетом двух мутаций на 25 маркерах, наш с Михаэлем общий предок родился задолго до Хейнриха, 550±390 лет назад, как я указал выше, то есть ориентировочно в интервале между 940 и 160 лет назад. Ну, на 430 лет назад я своих предков знаю, так что до того времени вряд ли. Хотя нельзя гарантировать, что кто-то из моих кровных родственников не попал в плен, и не унесло его в Германию. Либо, напротив, занесло моего прямого предка в 11-14 веках в Россию (вспомним на момент Ван-Дорна, он же Фандорин), чтобы в 1575 году в России родился Иван Клёсов, сын боярский, которому в 1636 году царским указом за рейтарскую службу дали надел в 300 гектаров на Курщине, и который дал жизнь - по прямой линии - сыну Кирею, тот – Сергею, тот – Кондрату, тот – Афанасию, тот – Парамону, тот – Федору, тот – Авксентию, тот – Василию, тот – Илье, тот - Ермолаю, тот - Ивану, тот - Алексею, а тот - Анатолию, стало быть, мне. Либо моего прямого предка тысячу лет назад или несколько позже занесло в Европу, и его (пра)n- внуком стал Хейнрих, а потом и Михаэль Хафман - сие мне пока неизвестно.
А на самом деле скорее всего было вовсе не так. Потому что все мы, с моей гаплогруппой – потомки одного рода ариев (R1a), и наш общий предок жил на территории нынешней России-Украины примерно 4900 лет назад. И предок наш отправился с сородичами-ариями походами и в Центральную Европу, и на юг, до Ближнего Востока, и в Иран, и в Индию. И везде разнес нашу, праславянскую (на Русской равнине) гаплогруппу и наши гаплотипы. И живут их потомки, а значит, члены нашего рода, и на Западе, и на Юге, и на Востоке. А род ариев – потому что именно этот род, точнее, его часть, пришел в Индию 3500 лет назад именно под именем ариев. И в Иран в те же времена – и опять под тем же именем. Через тысячу лет после того Дарий I, ахеменидский царь, выбил на барельефе в Накш-и-Рустаме: «Я, Дарий, великий царь, царь царей... арий, сын ария».
И дата та, 4900 лет назад, не с потолка взята, а из наших же гаплотипов (см. первый рассказ в этой книге). Потому что те мутации в них тикают как часы, с размеренной средней скоростью, что и позволяет вычислить то время, когда они в наших гаплотипах начали тикать. А именно, с нашего общего предка. Об этом речь в других рассказах в этой книге, и довольно подробно.
И вот к чему это приводит. Давайте посмотрим не на точные совпадения с моими 12 маркерами, а на те, которые отклоняются всего на один шаг. Всего на одну мутацию. Это – более информативное сопоставление, времени-то от нашего общего предка прошло немало, тысячи лет. Вот мутации и набежали. Так какая география моих сородичей-ариев? 26 человек.
Одну мутацию на моих 12 маркерах имеют 15 человек из России, по семи из Польши, Германии и Норвегии, шестеро из Финляндии, по четыре из Литвы, Англии-Ирландии, Швеции, двое из Пакистана. А также из Сирии и Шри-Ланки. Вот уже география походов ариев стала проясняться. А если по два отклонения, две мутации?
35 человек – Польша, 27 – Германия, 25 – Шотландия, 22 – Индия, 20 – Россия, 18 – Англия, 14 – Украина, 13 – Норвегия, по 10 – Греция и Италия.
Вот он, арийский ареал. Иной спросит – а Индия-то при чем? 22 человека – не шутка. Не случайность, таких случайностей не бывает. Так Индия – наши прямые родственники. 100 миллионов мужчин в Индии – наш народ, тот, которого в России больше половины, с гаплогруппой R1a. Их еще по старинке, а ныне политкорректно именуют «индоевропейцы». А на самом деле арии они, пусть потомки. Как и мы, славяне, R1a. Так наш древний род именуют в научных трудах.
На самом деле родственников у меня значительно больше, с теми же гаплотипами, но их потомки пока не сдавали ДНК на анализ.
А те, кто сдал, что могут видеть? А вот что. Взглянем опять на классический минимальный набор маркеров
DYS 19 - 388 - 390 - 391 - 392 - 393,
и на то, сколько повторов (то есть какая аллель) имеется в каждом маркере для разных групп людей. Напомним, что характерный рисунок этих повторов, в виде конкретных гаплотипов, определяет гаплогруппу, то есть генеалогическую общность людей, несущих одну потомственную характерную метку в ДНК.
Наиболее известный европейский гаплотип - так называемый "Атлантический модальный гаплотип" (АМГ), поскольку он характерен для многих людей, живущих вблизи Атлантического океана. Число повторов маркеров такое:
14 - 12 - 24 - 11 - 13 - 13
Более правильная, но более громоздкая запись была бы такой:
DYS19=14, DYS388=12, DYS390=24, DYS391=11, DYS392=13, DYS393=13.
Этот гаплотип принадлежит к гаплогруппе R1b.
У меня соответствующие шесть цифр следующие:
16 - 12 - 24 - 11 - 11 - 13
Как видно, я отклоняюсь от атлантического гаплотипа на четыре шага при шести маркерах. У меня - та же гаплогруппа R1, но другая подгруппа (R1a), но об этом ниже.
Продолжение истории в лицах. О президентах, царях и завоевателях
Томаса Джефферсона я уже здесь упоминал, показав два его маркера в числе повторов. Приведем его гаплотип, или "подпись", как иногда говорят генетики:
13 – 24 – 15 – 10 – X – Х – Х – 12 – Х – 12 – 15 – 27
Здесь - более длинная цепочка аллелей (повторов), чем я приводил выше, поскольку она записана в стандартной системе 12 маркеров. Но у Джефферсона (точнее, у его ныне живущих родственников) четыре из этих 12 не определялись, и я просто поставил вместо них ничего не значащие буквы. Эти стандартные 12 маркеров в правильном порядке следующие:
DYS# 383 - 390 - 19 - 391 – 385a – 385b - 426 – 388 – 439 – 389-1 – 392 – 389-2
У меня они такие:
13 – 24 – 16 – 11 – 11 – 15 – 12 – 12 – 10 – 13 – 11 – 30
Как видим, суммарно мой гаплотип по восьми маркерам отходит от гаплотипа Джефферсона на десять шагов. В среднем 1.25 мутаций на маркер. Это значит, что наш общий предок удален примерно на 50 тысяч лет (если считать правильно, с поправками). Его нет и близко.
Посмотрим на гаплотип Чингиз-хана (точнее, тот, который приписывают Чингиз-хану). Он для меня интересен вдвойне. Согласно молве, татаро-монголы, пройдя Русь огнем и мечом, сильно изменили генетический профиль русского народа, за исключением северных русских. Ну-ка, посмотрим, есть ли в моем гаплотипе монгол. Поскольку генеалогический рисунок Y-хромосомы передается только по мужской линии, всего один монгол за триста лет ига оставил бы неизгладимый след в своем потомстве. В общем-то я уже знаю путь моих прямых предков из Европы на Русскую равнину, а их братьев и племянников в Черноморские и Каспийские степи, и далее на восток, юго-восток и на юг, до Индийского океана, о чем расскажу позже, и в Монголию моих предков не заносило, но все-таки... Итак, гаплотип Чингиз-хана:
13 – 25 – 16 – 10 – 12 – 13 – 11 – 14 – 10 – 13 – 11 - 29
Отстоит от моего на 9 шагов, то есть в среднем 0.75 мутаций на маркер. Это – кака минимум 15 тысяч лет разницы, а на самом деле те же 50 тысяч лет. Но поскольку вопрос принципиальный, представим гаплотип хана в 25-маркерном варианте. Это – значительно более точная оценка:
13-25-16-10-12-13-11-14-10-13-11-29-18-8-8-11-12-26-14-22-27-11-11-12-16
Мой 25-маркерный гаплотип:
13-24-16-11-11-15-12-12-10-13-11-30-16-9-10-11-11-24-14-20-34-15-15-16-16
38 мутаций на 25 маркерах, 1.52 мутации на маркер. Еще дальше, чем от Джефферсона. Зашкаливает по всем критериям. То есть не то что далеко, а дальше некуда. Да и гаплогруппа другая, у меня R1a, у него С3. Мои с монголами предки, судя по всему, только из Африки вместе выходили, да и то, как недавно выяснилось, не было в Африке предков ни европейцев, ни монголов. Так что не повезло татаро-монголам с моими прабабушками, не дались они. Или молва преувеличивает масштабы тех событий.
Посмотрим на 12-маркерный гаплотип И. Сталина:
14 – 23 – 15 – 9 – 15 – 16 – 11 – 12 – 11 – 11 – 10 – 28
17 шагов в сторону от моего на 12 маркерах. 1.42 мутации на маркер. А на 25-маркерном гаплотипе Сталина -
14-23-15-9-15-16-11-12-11-11-10-28-17-9-9-11-11-25-16-21-28-13-13-14-14
39 отклонений. 1.56 мутаций на маркер. Оказалось, дальше есть куда, в сравнении с Чингиз-ханом. Да и гаплогруппа у Сталина другая, G2a. Это всё – и гаплотип, и гаплогруппу, определили у внука Сталина, который согласился помочь науке. Действительно, и то, и другое характерны для осетин.
Да и между Чингиз-ханом и Сталиным дистанция тоже огромна – 35 отклонений на 25-маркерных гаплотипах.
А вот гаплотип Пржевальского. Это тот, которого лошадь.
13 – 25 – 15 – 11 – 11 – 15 – 12 – 12 – 10 – 13 – 11 – 30
Вот это другой разговор. Сравните с моим:
13 – 24 – 16 – 11 – 11 – 15 – 12 – 12 – 10 – 13 – 11 – 30
Всего два отклонения на 12 маркерах. И понятно почему – мы с Н.М. Пржевальским принадлежим одному роду, R1a, к которому принадлежит большинство русских, и все восточные славяне, которых от 50 до 70% в старинных русских (а также украинских и белорусских) городах, городках, деревнях. Это отклонение соответствует примерно 2300 лет до общего нашего с ним предка, то есть близко к началу нашей эры. А общий предок нас, восточных славян России, гаплотипы которых известны, жил 4900 лет назад, как уже было упомянуто выше. Когда больше русских узнают свои гаплотипы, эта дата будет уточнена.
Пока рассмотрим интересный маркер, DYS388, второй по счету в «научной» записи, или восьмой в «стандартной». У "коренных" европейцев (обратите внимание на примеры выше) там обычно 12 повторов, как и у восточных славян. У выходцев с Ближнего Востока там часто 15 или 16 повторов (один из признаков гаплогруппы J1 или J2). Например, как в "Модальном гаплотипе коэнов" (МГК), «классическом» гаплотипе ближневосточных евреев и их современных потомков, как ашкенази, так и сефардов. В 6-маркерном варианте этот гаплотип выглядит так:
14 – 16 – 23 – 10 – 11 - 12
А в 12-маркерном расщепляется на два гаплотипа (маркеры, где произошло расщепление, выделены):
12-23-14-10-13-17-11-16-11-13-11-31
12-23-14-10-13-15-11-16-12-13-11-30
Прародитель этой генеалогической группы жил на Ближнем Востоке 4000 лет тому назад, а оба «гаплотипа коэнов» возникли уже в нашей эре, один примерно 1650 лет назад, другой – 1000 лет назад. Первый (верхний) повел действительно священник, коэн, второй – похоже, выживший в 7-м веке нашей эры прямой потомок царя Давида. Коэном по рождению он не был, но принял этот титул согласно традиции, как уважаемый человек при отсутствии в сообществе «легитимных» коэнов. Так получается, хотя надо, конечно, уточнять.
Перейдем к еврейской тематике.
О евреях и палестинских арабах
Кстати, о модальном гаплотипе коэнов
14 - 16 - 23 - 10 - 11 - 12
Он является действительно частым среди еврейских священников, коэнов, и в целом характерен среди евреев ближневосточного происхождения. Этот гаплотип практически отсутствует среди практически всех других народностей (кроме арабов), и что любопытно – его очень мало среди палестинских арабов, да и то, что есть – случайности мутаций. Но зато его много у бедуинов, и появился он как минимум 9 тысяч лет назад. Так что Авраам, или как его звали на самом деле, общий патриарх евреев и арабов, его от бедуинов унаследовал, и передал по цепочке поколений и евреям, и арабам. И, естественно, среди евреев - коэнам, через своего потомка Аарона, брата Моисея.
Термин «модальный гаплотип» часто вносит путаницу. Обычно «модальным» гаплотипов называют тот, что наиболее часто встречается той или иной популяции. Правильно было бы говорить «предковый» гаплотип, но его мы часто не знаем. Поэтому в ДНК-генеалогии принят термин «базовый гаплотип», то есть максимально приближенный к предковому, или сам предковый, когда это выяснено. А модальным гаплотипом можно назвать тот, который наиболее часто встречается, например, в Москве. Он вообще может оказаться китайским или азербайджанским. Шутка, конечно, но в каждой шутке есть доля правды.
А вот и «модальный» гаплотип палестинских арабов:
14 - 17 - 22 - 11 - 11 - 12
Три шага в сторону от коэнов всего на шести маркерах, что значит – примерно пять тысяч лет от общего предка. А на более протяженных гаплотипах получается более точно, четыре тысячи лет до общего предка евреев и арабов. Случайно или нет – это время жизни библейского Авраама, как следует из анализа библейских текстов. Так что легенда-легендой, но основания у нее могут быть. Как я упомянул, не обязательно общего предка евреев и арабов звали Авраам, но такой определенно был, четыре тысячи лет назад.
А как насчет бедуинов? У них два «модальных» гаплотипа
14 - 15 - 23 - 10 - 11 - 13
14 - 16 - 23 - 10 - 11 - 12
Второй – «модальный гаплотип коэнов». Мы видим всю абсурдность этого названия. У первого - два шага в сторону от первого на шести маркерах. Ближе, чем палестинцы, но все равно 3200 лет от общего предка. Три мутации дали бы пять тысяч лет. Вот и погрешность определения на таких коротких гаплотипах. На самом деле – те же 4 тысячи лет, с соответствующей погрешностью.
Ну ладно, вот курдские мусульмане:
14 - 15 - 23 - 10 - 11 - 12
Один шаг в сторону. Уже теплее. Но совпадения все равно нет.
Раз уж мы заговорили о евреях, то заметим, что среди евреев-мужчин выделяются несколько групп, которые оставили свой ранний след в ДНК-генеалогии – коэны (наследственные высшие священники), левиты (другая группа наследственных служителей обрядов) и израэлиты, к которым принадлежат 90% евреев, в том числе ашкенази (европейские евреи), сефарды (северо-африканские и пиренейские евреи) и остальные евреи. Эфиопские евреи и народность Лемба в Южной Африке, которые себя причисляли к евреям, и о чем они – по их словам – знали с незапамятных времен, евреями по генеалогическим корням совсем не оказались. Первые – просто эфиопы, как и все эфиопы, вторые – арабы, йеменцы.
Так как насчет моей гаплогруппы, R1a? С одной стороны, мои ДНК-генеалогические корни в Европе, с другой стороны – есть некоторые сходства с западно- и восточно-европейскими гаплотипами. Куда я ближе? И сколько вообще гаплогрупп?
История гаплогруппы R1a
Эта история подробно рассказана в первых двух главах этой книги, и в последующих, так что не буду повторяться. Если коротко напомнить – то мои предки появились в составе так называемой бета-гаплогруппы, или сводной гаплогруппы ВТ, из которой и отделились все гаплогруппы от В до Т, на протяжении как минимум 30 тысяч лет. А сама бета-гаплогруппа появилась примерно 65 тысяч лет назад. По мере расхождения людей по разным направлениям, формировались рода, которые и есть гаплогруппы. Мой будущий род который тогда был гаплогруппой К, отошел от сводной гаплогруппы IJK, и направился за лучшей долей на восток, тогда как род I ушел на запад, в Европу, а род J – на юг, в Месопотамию. Из этой диспозиции можно предположить, что расходились они из из Европы, или с Русской равнины. Скептики могут заголосить, что вот, мол, опять Россия на первом месте, но на то они и скептики. Пусть попробуют сами выдвинуть наименее противоречивую историю исходных миграций гаплогрупп, но на это надеяться не приходиться. Скептики сами ничего не выдвигают. На то они и скептики.
Так вот, по пути на восток от гаплогруппы К отошли рода NO и P, причем гаплогруппа Р, видимо, уже образовалась в Южной Сибири. От нее отделились гаплогруппы Q и R, это – тоже европеоидные группы, как и IJK, и поначалу NO, и Р. По-каким-то причинам, пока неясным, N и Q стали в значительной степени монголоидными, О стала восточно-азиатской, а Р осталась европеоидной, как и образовавшиееся из нее гаплогруппы R, и затем R1, R1a, R1b. Два последних рода совершили грандиозный по масштабам поход в Европу, первый по южной географической дуге - через Тибет, Индостан, Иранское плато, Анатолию, и прибыли на Балканы 10-9 тысяч лет назад, вторые – по северной дуге, через Южный Урал, северный Казахстан, среднюю Волгу, и далее, разойдясь по разным направлениям, взяли Европу в клещи между 4800 и 4500 лет назад, и начали агрессивное расселение. Практически все гаплогруппы «Старой Европы» или исчезли, или бежали в разных направлениях. Бежала (или организованно отошла) на восток и гаплогруппа R1a, прибыв на Русскую равнину примерно 4900 лет назад. Это были арии, которые уже через несколько веков разойдутся с Русской равнины на восток, откуда частью двинутся в Индию, как индоарии, частью в Зауралье, вплоть до Алтая и Китая, а также на юго-восток, через Среднюю Азию на Иранское плато, как авестийские арии, и на Ближний Восток, как митаннийские арии.
А остальные носители гаплогруппы R1a останутся жить на Русской равнине, оставив там многие археологические культуры, и в 1-м тыс до н.э., 3000-2000 тысячи лет назад, отправятся опять перезаселять Европу. Потому почти все гаплотипы R1a в Европе – с Русской равнины, праславянские. В итоге большинстве европейских ветвей гаплогруппы R1a присутствуют и в нынешней Росссийской Федерации, а доля гаплогруппы R1a в РФ достигает 63%, а местами и выше, да и не везде измеряли.
Вот почему огромная «индоевропейская полоса» гаплогрупп R1a, если использовать лингвистический термин, протянулась от запада Европы до Индии и Ирана. На самом деле это наложение многих миграций гаплогруппы R1a – сначала древних миграций из Европы на Равнину, затем с Равнины по разным направлениям, включая в Индию, и затем опять с Равнины – уже в Европу. Так Европа перешла на единую языковую семью – арийскую, которую потом по политическим соображениям назвали индо-европейской. Вот и связка между Европой и Индией, которую проложила гаплогруппа R1a.
Таким образом, на Балканах, и далее на Русской равнине, мои прямые предки говорили на языке, который положил начало индоевропейской семье языков, включающей английский, французский, немецкий, русский, испанский, греческий, несколько индийских языков, таких как бенгали и хинду, многочисленную группу иранских языков, и много других. Правда, по иронии судьбы, а точнее, по прихоти (или по незнанию) лингвистов язык моих прямых предков назвали «иранским», только потому, что «иранские» языки лингвисты изучали более детально, и не имели понятия, что этот язык на самом деле принесен на Иранское нагорье моими прямыми предками, которые называли себя ариями. Под этим именем они и пришли в Индию и Иран.
По справедливости, это иранские языки следовало было назвать «арийскими», и тогда не было бы таких ситуаций, когда живших на Днепре, на Дону, в Причерноморье современные историки, лингвисты, археологи считают говорившими на «иранских языках», как «иранцами» именуют скифов и сарматов, которые отродясь в Иране не были. По той же причине жителей уральского Аркаима (3800-3600 лет назад) именуют «индоиранцами», хотя у ариев, жителей Аркаима того времени ни Индии, ни Ирана и в проекте не было. Но консерватизм и косность – вещи упрямые, и лингвисты и археологи продолжают держаться за эти давно устаревшие термины, оправдываясь, что они ни Индию, ни Иран не имеют в виду, а «так принято». В смысле, так когда-то назвали, не зная истинных причин, так и провелось, что наши предки были якобы «иранцами». Не любят почему-то наши историки и лингвисты ничего славянского до нашей эры, видимо, до смерти боятся, что шовинистами назовут. Пусть уж лучше славяне будут «иранцами», так спокойнее.
Как бы там ни было, а сотни миллионов людей, живущих в Европе, особенно на севере Франции и в Англии, Шотландии, Германии, Швеции, в Прибалтике, Украине и Белоруссии, в Индии и Иране, на Ближнем Востоке и Аравийском полуострове, и в России вплоть до Сибири и Дальнего Востока, являются потомками этой гаплогруппы R1a. Моей гаплогруппы. Нашей, славянской, если говорить о территориях, где говорят на славянских языках.
А часть сводной гаплогруппы Р, как я уже рассказывал, из Южной Сибири стала гаплогруппой Q и ушла дальше на север, в ледники, в итоге стала эскимосами, часть посуху перешли на Аляску и стали американскими индейцами. Но у них были уже другие субклады-маркеры.
Пошли бы мои прямые предки дальше на северо-восток - быть бы мне эскимосом. Перебрались бы через пролив с Аляской - быть бы мне американским индейцем. Пошли бы южнее - быть бы мне индусом, китайцем, а то и полинезийцем.
Не судьба...
CE – ЧЕЛОВЕК
ЧАСТЬ ВТОРАЯ – О ЖЕНЩИНАХ
В отличие от мужчин, у женщин нет Y-хромосомы. Женская секс-хромосомная пара состоит из ХХ хромосом. Мужская – из XY хромосом. Сперматозоид равновероятно несет только одну – либо X, либо Y хромосому. Проскочит в яйцеклетку Х – быть девочке. Проскочит Y – стало быть, мальчик, с его возможностями генеалогического анализа Y-хромосомы, как описано в предыдущей главе.
А как быть женщинам? Может, у них можно Х-хромосому так же анализировать?
Нельзя. Потому что Х-хромосомы у женщин перетасовываются. Одна получена от мамы, вторая – от папы. Поди разберись, какие там предки и от кого.
В любом случае, к женщинам опять нужен свой подход. И природа его предоставила.
Помимо ДНК хромосом, которые хранятся в ядре каждой клетки, молекулы ДНК находятся также в митохондриях. Митохондрии – это маленькие образования, плавающие во внутриклеточной жидкости, в цитоплазме. Их – от нескольких сотен до нескольких тысяч, даже до десятков тысяч на каждую клетку. И в каждой – короткая молекула ДНК, в виде несимметричного кольца. Длина ее – всего 16 с половиной тысяч нуклеотидов. Сравните с мужской хромосомой Y, которая в три тысячи раз длиннее, 50 миллионов нуклеотидов.
Митохондриальная ДНК (мтДНК) состоит из двух частей – выпирающая в сторону петля, и остаток кольца. Оказалось, эта петля является носителем генеалогической информации, не хуже, чем у мужчин, но совершенно по другому.
В мтДНК нет таких тандемных повторов, как в Y-хромосоме. Там нет подобных маркеров, о которых шел рассказ в первой части. Но мутации – есть. Время от времени, причем намного реже, чем у мужчин, считывающий фермент ошибается и вместо одного нуклеотида вставляет другой. Или вообще вставляет лишний. Поэтому мутации записываются, например, так – 1651С. Поскольку известно, что в «стандартной» мтДНК нуклеотид под номером 1651 – тимин (Т), сразу ясно, что в этом положении тимин заменен на С (цитозин). Или запись такая: 315.1С. Это значит, что после 315-го нуклеотида в «стандартную» цепь вставлен один лишний цитозин.
Иначе говоря, у мужчин ДНК-генеалогия основана на изменении числа повторов определенных маркеров в хромосомной ДНК, а у женщин – на разовых нарушениях одиночных нуклеотидов в митохондриальной ДНК. То есть совершенно другой принцип. Маркеров как таковых у женщин нет, вся петля ДНК – один сплошной маркер. А сравнивают – со «стандартной» мтДНК.
Поскольку мтДНК в основном некодирующая, то эти мутации в петле ни к чему жизненно важному не приводят. Просто запись в генетической книге учета.
А что такое «стандартная» мтДНК? С чем сравнивают-то?
А это так получилось. В 1981 году генетики в нашем Кембридже, заречном пригороде Бостона, были готовы провести первое определение последовательности, или «первичной структуры» мтДНК. Нужна была любая плацента, клетки которой крайне богаты митохондриями. А в соседнем госпитале как раз рожала женщина. Взяли ее плаценту, выделили митохондрии, оттуда – мтДНК, и провели полный анализ ее последовательности. Поскольку это была первая последовательность мтДНК– её и взяли за международный стандарт. И мутации в последовательности отсчитывают от нее.
Впоследствии оказалось, что эта стандартная мтДНК присуща именно европейскому типу, и мтДНК женшин с «европейскими корнями» отклоняются от нее всего на несколько позиций, или вообще не отклоняются. Фамилия этой кембриджской женщины осталась нераскрытой. А последовательность ее мтДНК в генетике называют «кембриджской стандартной последовательностью».
Здесь – важное отступление. Мужчины получают свои митохондрии от мамы, но своим сыновьям не передают. Поэтому митохондриальная ДНК на каждом мужчине терминируется. Нет девочек в роду – мтДНК терминировалась на мальчиках, связь этой линии с праматерью («митохондриальная Ева») потерялась. Нет мальчиков в роду – терминировалась Y-хромосома, потерялась генеалогическая связь с праотцом («хромосомный Адам»). Нет детей – полная терминация генеалогической информации от отца с матерью. Но каждый мужчина имеет мтДНК, и ее анализ дает такую же генеалогическую информацию, как и анализ мтДНК его матери или сестры. А, повторяю, наличие такой информации – генеалогический «прострел» к прародителям - десятки, а то и больше сотни тысяч лет назад, как к «Еве» (мтДНК), так и к «Адаму» (Y-хромосома), плюс информационное богатство всех мутаций на историческом пути.
Здесь надо опять повторить, что «митохондриальная Ева» - это вовсе не первая женщина, а ближайшая по времени прародительница всех женщин на Земле. Та, к которой сходятся генеалогические нити от всех живущих на планете. Ее мама – не ближайший прародитель, если у мамы была только одна дочь – «Ева». Так что у «Евы» еще один ограничительный признак – у нее должно было быть по меньшей мере две дочери. Чтобы от Евы и пошел тот генеалогический «разбег», в итоге породивший все человечество. Подруги Евы не стали прародителями человеческого рода, а также не стали те, кто жили вокруг, или в отдалении, тысячи и десятки тысяч, а то и сотни тысяч лет до «Евы». Их потомство не оставило генеалогических следов в живущих в настоящее время на Земле.
Естественно, я провел анализ также своей мтДНК. Точнее, провели для меня, но это дело не меняет. Результат был дан в первой части этого рассказа. Повторю:
16519С 263G, 309.1C, 315. 1C, 477C
Поскольку мтДНК – кольцо, то неважно, откуда начинать считать. Договорились считать от середины выпирающей петли как наиболее заметного образования. А поскольку в «стандартной» мтДНК всего 16569 нуклеотидов, то моя мутация (точнее, моих бабушек) номер 16519С произошла недалеко от точки отсчета, на 50 нуклеотидов ниже начала. Там, от номера 16001 и до 16569, находится область, относительно богатая мутациями. Ее называют областью низкого разрешения, или HVR1 (hypervariable region 1). Вторая, область высокого разрешения, HVR2, находится сразу за первой, с нуклеотида 1 до 580. Собственно, часто только там, в этих двух областях, и проводится генеалогический анализ митохондриальной ДНК.
Часто, впрочем, «высоким разрешением» в ДНК-генеалогии называют полное совпадение мутаций в обеих областях, HVR1 и HVR2, у двух человек. Тогда с высокой вероятностью оба имеют общую «бабушку» на протяжении последних примерно 1000 лет.
Итак, верхняя строка в «моих» мутациях относится к HVR1, нижняя – к HVR2. Более детально это означает – одна мутация с заменой тимина в позиции 16519 на цитозин, и четыре мутации в позициях 263 (аденин на гуанин), 309.1 (вставка лишнего цитозина), 315.1 (еще одна вставка цитозина), и 477 (замена тимина на цитозин).
Строго говоря, и мутациями-то эти изменения назвать нельзя. Это – просто отклонения от «стандартной», условно выбранной последовательности. То есть вполне может быть, что это не у меня мутация в виде замены тимина на цитозин в позиции 16519, а у той женщины, чья мтДНК принята за стандарт, древний цитозин в этой позиции мутировал на тимин. Все в мире относительно. Но для наших целей генеалогического анализа это не так важно.
В любом случае, мой мтДНК гаплотип, или система отклонений от «стандартной мтДНК», относится к гаплогруппе H, или Helena, одной из семи европейских гаплогрупп. Она появилась в Европе 20 тысяч лет назад, по некоторым оценкам. Но об этом чуть позже. А сейчас напомним, что такое гаплотипы и гаплогруппы, чтобы не путаться.
Гаплотип – это ваша индивидуальная генетическая характеристика. При рассмотрении мтДНК – это ваш набор отклонений от «кембриджской стандартной последовательности», например, в виде двух строк, что приведены выше (для HVR1 и HVR2, соответственно). А гаплогруппа – это генетическая характеристика определенной общности людей, которые имели одну общую «пра»бабушку, более недавнюю, чем «митохондриальная Ева». Их древние предки часто передвигались в одной группе в ходе миграций. Гаплогруппа показывает, к какой генеалогической ветви человечества вы относитесь. Их обозначают буквами алфавита, от А до Z, плюс многочисленные подгруппы.
Например, европейские гаплогруппы – H, J, K, T, U, V, X. Ближневосточные – N и M. Азиатские – A, B, C, D, F, G, M, Y, Z. Африканские – L1, L2, L3 и M1. Полинезийская – B. Американские индейцы – А, B, C, D, и редко Х. В последнее время к европейским гаплогруппам добавили N1, U4, U5 и W. Но надо помнить, что митохондриальные, женские гаплогруппы не имеют ничего общего с хромосомными, мужскими гаплогруппами. Индексы у них тоже разные.
Недавно провели анализ древнейших костей человека на Русской равнине, с датировкой 30-40 тысяч лет назад, и оказалось, что он имел гаплогруппу U2. И знаете, где этой гаплогруппы сейчас больше всего? Во Франции и в Индии, в этой последовательности.
Выше было упомянуто, что гаплогруппа Н появилась 20 тысяч лет назад. И вот здесь надо сделать замечание. Если расчетный аппарат при работе с гаплотипами Y-хромосомы хорошо отлажен, то с мтДНК с этим значительно хуже. Вернемся опять к моей мтДНК:
16519С
263G, 309.1C, 315.1C, 477C
Мало того, что всего пять «точек опоры», по сравнению с 67, а то и 111 в мужских гаплотипах, так там и мутации происходят раз в три с лишним тысячи лет. Так что с генеалогическими расчетами не разбежишься. Еще одна особенность мтДНК – это то, что они весьма хаотически разбросаны по территориям, например, по всей Евразии. Дело в том, что по исторической традиции – где узаконенной, а где так сложилось – жен и наложниц умыкали где получится. Помимо того, если даже не умыкали, то жены обычно приходили (и сейчас приходят) в род, племя мужа, а не наоборот. Это и привело к фактическому рассеянию мтДНК по территориям, так что о направленных миграциях мтДНК, в отличие от Y-ДНК, говорить не приходится. Бывают, впрочем, случаи, когда мтДНК образуют свои «оазисы», но это случаи довольно редкие и экзотические, как правило, на изолированных территориях, с затрудненными контактами с внешним миром на протяжении тысячелетий, например, среди некоторых племен американских индейцев. Тогда это очень интересно и информативно, но это скорее исключение, чем правило. И опять, с одной мутацией на несколько тысяч лет не до ДНК-генеалогии. Обычно при этом просто фиксируется территория, на которой такой «оазис» мтДНК найден. Этим занимается обычно наука популяционная генетика, не ДНК-генеалогия.
Вот несколько примеров, что такой анализ дает. Жена последнего русского царя Николая II, Алиса Гессен-Дармштадтская, внучка английской королевы Виктории, имела такой гаплотип:
16111Т, 16357С, 16519С
263G, 315.1C, 524.1A, 524.2C
У нее с королевой Викторией точно такие же мутации в зоне высокого разрешения, как и у моих бабушек. И гаплогруппа та же, H, но это уже индексы митохондриальных родов, или гаплогрупп. Так что европейская общая бабушка у нас с ними одна. Кстати, принц Филип, дюк Эдинбургский и муж ныне здравствующей королевы Елизаветы II, имеет точно такой же гаплотип, как и Алиса. Что не удивительно, они – племянники.
Поехали дальше. Как насчет Марии Антуанетты? Королевы Франции, обезглавленной в 1793 году в возрасте 37 лет? Жены Людовика XVI? Вот ее гаплотип:
16519С 152С, 194T, 263G, 315.1C
Ничего не напоминает? Ну как же, гаплотип моих бабушек по материнской линии:
16519С 263G, 309.1C, 315. 1C, 477C
Та же «входная» мутация в подгруппу гаплогруппы Н, тот же вариант петли мтДНК низкого разрешения, те же две мутации из четырех в области высокого разрешения. Даже мутация 315.1С та же, вставка лишнего цитозина в том же месте. Определенно общая бабушка у Марии Антуанетты с моими, причем уже в Европе, относительно недавно. А как недавно? Да, видимо, несколько тысяч лет назад. А «недавно» - потому что в мтДНК много родов, или гаплогрупп, и то, что мы с королевой Викторией или Марией Антуанеттой входим в одну гаплогруппу, уже означает относительно близкое родство. Принципиальное, если угодно. Один род.
Раз уж мы погрузились в рассмотрение мтДНК, посмотрим на генеалогическое древо английской королевы Виктории, которое доходит до семьи Николая Романова, то есть дочерей Николая II и Александры, погибших в с родителями в 1918-м, а также до принца Филипа по параллельной линии. Как мы уже рассказывали, мтДНК передается в поколениях исключительно по женской линии. Потомству мтДНК передает только мать, из своей яйцеклетки (женской половой клетки). Мужчина мтДНК не передает. Не мужское это дело.
Вот как выглядит женская линия королевы Виктории до Александры Федоровны Романовой, она же Алиса (Аликс) Гессен-Дармштадская, внучка английской королевы:
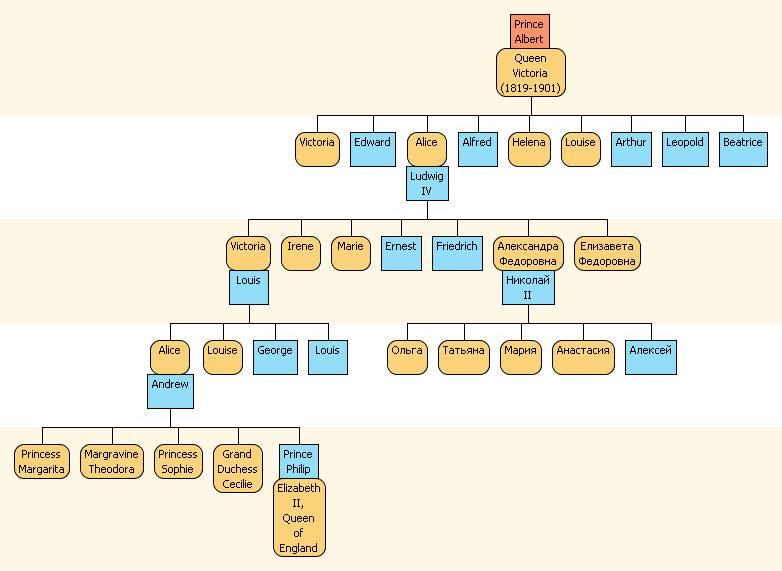
Рис. 046. Генеалогическое дерево (женская линия) от королевы Виктории (род. 1819 год) до ныне здравствующей английской королевы Елизаветы II и ее сестер, и до последней царской семьи России.
На этой схеме показано, как мтДНК переходит по желтым, женским линиям от королевы Виктории через ее дочь, великую княгиню Алис, и далее расходится по нескольким параллельным потокам, из которых здесь показаны два – до герцога Эдинбургского Филипа, мужа нынешней королевы Великобритании Елизаветы II, и его четырех сестер, с одной стороны, и до Александры Федоровны Романовой и ее детей – княжен Ольги, Татьяны, Марии, Анастасии и царевича Алексея. Таким образом, все мужчины и женщины, показанные на этой генеалогической схеме (кроме супругов, пришедших «со стороны»), имеют одну и ту же мтДНК. Кстати, её для тестирования и определения картины мутаций в мтДНК членов царской семьи во время расследования, чьи же останки найдены у Екатеринбурга, предоставили несколько членов монарших семей Европы, для схемы выше – принц Филип.
Но по этой схеме Николай II выпадает из рассмотрения. Он – муж Александры Федоровны, пришел «со стороны». Поэтому посмотрим на другую схему, фрагмент генеалогического дерева ветви европейских монархов, идущей по женской линии от Луизы фон Гессен-Кассель, бабушки Николая Второго. Мать самой Луизы была Шарлотта Датская, бабушка – София Фредерика, супруга принца Датского и Норвежского.
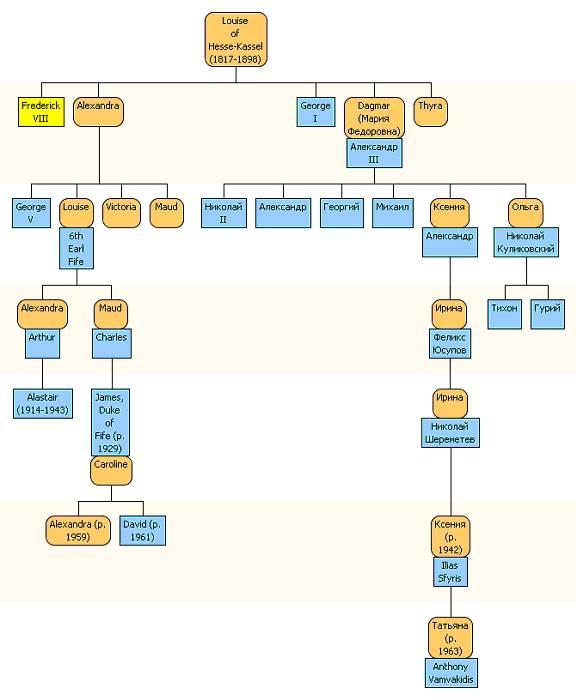
Рис. 047. Генеалогическое дерево (женская линия) от Луизы фон Гессен-Кассель (1817 г.р.), бабушки русского царя Николая II, до ее современных потомков, включая потомков русского царя Александра III
Эта ветвь, естественно, неполная, иначе в ней должны были быть отражены десятки, если не сотни человек. Суть в том, что здесь показаны две отдельные ветви, потомки которой предоставили свою мтДНК для идентификации мтДНК Николая II, и для сопоставления с ДНК других останков из могилы при известном расследовании. ДНК для анализа предоставили три человека – (а) принц Филип, муж ныне здравствующей английской королевы Елизаветы, (б) Ксения Николаевна, потомок родной сестры Николая Второго, Ксении Александровны, и английский герцог Джеймс Карнеги, потомок королевского рода по материнской линии.
Если мы проследим по женской линии, то есть по светлым фигуркам от королевы Виктории вниз по поколениям (первая генеалогическая схема выше), мы увидим, что принц Филип должен иметь ту же мтДНК, что и дети Александры Федоровны, включая Анастасию и царевича Алексея. Как мы помним, мтДНК передается мужчинам в первом поколении, а затем на них обрывается. А вот у Николая Второго мтДНК будет уже другая, он в первой генеалогической схеме «со стороны», его мтДНК - по его матери и бабушек по материнской линии.
Зато согласно второй схеме, мтДНК Николая Второго будет та же, что у его бабушки Луизы по материнской линии, и та же ДНК будет и у его сестры Ксении, и у ее правнучки Ксении, что родилась в 1942 году, и у герцога Джеймса Карнеги, который родился в 1929 году и до сих пор здравствует.
И вот именно таким образом было показано, что останки, найденные в могилах, принадлежат именно царской семье. Там, правда, все время поднимаются новые вопросы, но для целей этого очерка они не так важны. В любом расследовании есть вопросы. Было бы желание.
мтДНК И ГЕНЕАЛОГИЧЕСКИЕ БАБУШКИ
Итак, у меня, а на самом деле у моих мам и бабушек, гаплогруппа H. Их, подгрупп гаплогруппы Н, насчитывают несколько десятков вариантов. На мой вариант сразу указывает «входная» мутация 16519С.
Естественно, я попытался найти, у кого такие же мутации в мтДНК, как у моих бабушек по прямой материнской линии. Оказалось, и уже давно, что «входную» мутацию 16519С имеют 5739 человек из базы данных, и их количество растет в базе день ото дня. Кстати, в первой версии этого рассказа, написанной несколько лет назад, эту «входную» мутацию в той же базе данных имели 2194 человек, и с тех пор в день в среднем каждый день их количество нарастало на 5 человек. Так что «день ото дня» - это вовсе не преувеличение. Стало понятно, почему эта петля – «низкого разрешения». Введение в поиск четырех других мутаций из HVR2 резко сбросило число родственников – до 22 человек. Несколько лет назад их было семь.
По запросу генеалогических организаций, люди, входящие в базу данных, предоставляют сведения о своих родственниках. Это – если они хотят получить доступ к такой же информации от других. Многие предоставляют данные в виде целых генеалогических деревьев. Таким образом, можно взглянуть, а кто же те женщины, жившие сотни лет назад, у которых мтДНК полностью совпадает с моей, то есть с моими прямыми бабушками по материнской линии. А значит, с кем у меня есть прямая генеалогическая связь по материнской линии. С теми, у кого мтДНК имеет мутации 16519С, 263G, 309.1C, 315. 1C, 477C. Вот имена тех, кто предоставил сведения о своих предках:
Margaret Stayse-Dewey, родилась в 1549 году в Wyndmonham, Norfolk, East Anglia, England
Elizabeth Bailey, родилась в Англии в 1782 году
Teresa Farkas, родилась в Венгрии в 1850 году
Elizabeth Bailey (уже другая), родилась в Честере, Южная Каролина, в 1775 году, году основания США
Fredrika Heinemann, родилась в Нью-Йорке, США, в 1846 году
Lucinda Pope, родилась в 1852 году, страна рождения неизвестна
Margaret Criswell, 1810 года рождения, Оксфорд, Ирландия
Szymanska, 1810 года рождения, Польша.
Angelique Tirad, родилась в 1793 году в США, штат Миссури.
Это только начало, там уже в списке 301 человек. А еще недавно были только 22 человека...
Когда у меня с ними была общая бабушка – оценить трудно. Мутации мтДНК – редкие. При полном совпадении мутаций в мтДНК общая бабушка могла жить от нескольких тысяч до десятков тысяч лет назад.
А что значит – мутации редкие? По оценкам генетиков, мутации в мтДНК происходят со скоростью 1/100000 на нуклеотид на поколение. Как я приводил выше, генеалогический анализ проводят на 569 нуклеотидах области HVR1 и 580 нуклеотидах HVR2, всего на 1149 нуклеотидах. То есть одна мутация происходит там в среднем на 100 поколений, или на 2500 лет. Но было доказано, что в этих областях мутации происходят более часто, чем в нуклеотидах мтДНК в целом. Ну, например, одна мутация – примерно на тысячу лет. Я был бы рад привести более точные цифры, но среди специалистов данные сильно расходятся.
Принято считать, что при полном совпадении зоны «низкого разрешения» HVR1 с такой же зоной в мтДНК другого человека, имеется 50%-ная вероятность того, что их общая «бабушка» жила в течение последних 52 поколений, или 1300 лет. И то, если гаплогруппа одинаковая. Если же при этом гаплогруппы разные, то совпадение случайное, вызванное «конвергентной эволюцией», и общая «бабушка» может отстоять во времени на десятки тысяч лет. А при полном совпадении «высокого разрешения», на обеих зонах HVR1 и HVR2, есть 50%-ная вероятность того, что общая «бабушка» жила на протяжении 28 поколений, или 700 лет. Об этом я уже упоминал.
Теперь взглянем на мтДНК людей известных. Ну, например, на Николая II Романова. Он получил свою мтДНК, естественно, от матери, Марии Софии Фредерики Дагмары (Марии Федоровны), а та, в свою очередь, от своей матери, королевы Дании Луизы Вильгельмины Фредерики Каролины Гессен-Кассельской:
16126С, 16169(С,Т), 16294T, 16296T
73G, 263G, 315.1C
Напомню мой гаплотип:
16519С
263G, 309.1C, 315. 1C, 477C
Мы сильно расходимся с королевой Дании в области низкого разрешения, но имеем две одинаковые мутации в области высокого разрешения. В целом, родство весьма отдаленное. И действительно, у моих бабушек гаплогруппа Н, у королевы Дании и Николая II – гаплогруппа Т. Кстати, племянник Николая II по материнской линии, князь Николай Трубецкой, а также родной брат Николая II, великий князь Георгий Александрович Романов имели совершенно такой же гаплотип, как и Николай II. Что вполне естественно, так и должно быть, поскольку линия - материнская.
Я не буду здесь пояснять, почему у второй мутации в области низкого разрешения у Николая II (и Трубецкого, и Георгия Романова) оказалось одновременно два нуклеодида (С,Т). Просто упомяну – «гетероплазмия».
Про жену Николая II, Алису Гессен-Дармштадтская, я уже писал выше, как и про Марию Антуанетту.
Чтобы у читателя не осталось впечатления, что все мтДНК имеют похожие мутации, приведу несколько других примеров знаменитых гаплотипов. Три из них относятся к останкам древних людей, найденных в мумифицированном виде, во льдах. Один – «Ледовый человек», Отци, живший 5300 лет назад:
16224С, 16311С (гаплогруппа К1).
Этот конкретный гаплотип именно в таком виде был найден всего у одного ныне живущего человека из примерно 60 тысяч тестированных на мтДНК, по крайней мере на сегодня. Его имя – Daniel Crain. Его наиболее удаленная прабабушка – Ann Wilmoth, родилась в 1866 году. Варианты этого гаплотипа имеются у пары сотен человек в базе данных, но в основном с включением, видимо, более поздней мутации 16519С («моей» мутации).
Другой «снежный человек» – Чеддармен, Чеддарский человек, живший 9000 лет назад:
16192Т, 16270Т (гаплогруппа U5a)
Третий, точнее, третья – «Хуанита», инка из Перу, жившая 500 лет назад:
16111Т, 16223Т, 16290Т, 16319А (гаплогруппа А).
Наконец, Лука-евангелист, умерший в Греции в 150 г, в возрасте 84 года. То, что это именно Лука – интерпретация, соображение, но гаплотип настоящий:
16235G, 16291T (гаплогруппа Н).
Поскольку гаплогруппа Н уже встречалась нам чаще всего, остановимся на ней более подробно. Хотя бы потому, что это гаплогруппа моих бабушек, и моя тоже.
Эта европейская гаплогруппа возникла при заселении Европы 20-40 тысяч лет тому. Вот как это было.
ЗАСЕЛЕНИЕ ЕВРОПЫ
Говоря генеалогически, были две основные волны прибытия древних «иммигрантов» в Европу. Первая – не менее 45-50 тысяч лет назад, когда прибыли первые поселенцы, но откуда – наука не знает, но предполагает, что или с востока, с Русской равнины, или с юга, со стороны Месопотамии. Некоторые говорят, что из Африки, но данных ни у кого нет. Они еще застали в Европе неандертальцев, по крайней мере, застали их первые переселенцы, которых относят к кроманьонской группе.
Неандертальцы с приходом наших предков быстро вымерли, отчего – неизвестно. Следов боев неандертальцев с нашими предками не обнаружено, похоже, этих боев просто не было. Имеется много древних пещерных наскальных изображений охоты, но ни одного – сражений с неандертальцами. Детей неандертальцы не оставили, их генетическая линия полностью терминировалась. Потомства неандертальцев с нашими предками тоже не было, судя по всему, хотя трудно себе представить, чтобы за тысячи лет совместной жизни (по крайней мере, территориально) наш предок не соблазнился бы неандертальской дамой. Или наоборот. Скорее всего, дело было более серьезно, скажем, хромосомы наших предков были несовместимы с неандертальскими. Например, на наши 46 хромосом у тех было 44 или 48 (как, например, у шимпанзе), так что хромосомные пары не могли образоваться, и потомства просто быть не могло. Или могло, но своего потомства не давало. Типа как у мула, дитя любви лошади и осла, извините за сравнение. Так или иначе, неандертальцы – не наши предки.
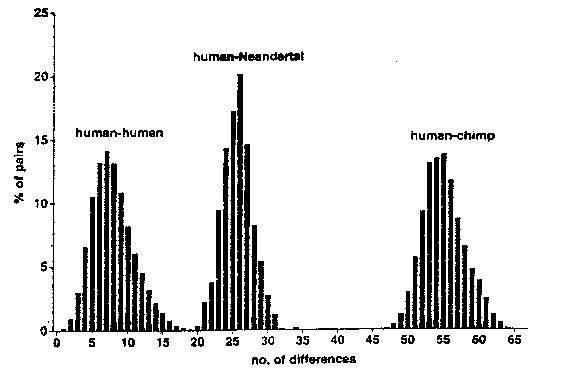
Рис. 048. Иллюстрация различий в последовательности мтДНК между разными людьми (первая кривая – среднее различие в 8±3 нуклеотидов), людьми и неандертальцами (вторая кривая – среднее различие в 27±2 нуклеотидов), и, для сравнения, людей и шимпанзе (третья кривая – среднее различие в 55 нуклеотидов).
А вот и одно из доказательств, что не наши предки. Эта диаграмма позаимствована из научной статьи Крингса с соавторами, и показывает различия в последовательностях мтДНК между разными людьми (первая кривая – среднее различие в 8±3 нуклеотидов), людьми и неандертальцами (вторая кривая – среднее различие в 27±2 нуклеотидов), и, для сравнения, людей и шимпанзе (третья кривая – среднее различие в 55 нуклеотидов).
Генетический анализ другого неандертальца, найденного на расстоянии 2500 км от первого, показал сходные результаты. Ясно, что неандертальцы не имеют прямого генеалогического отношения к людям. Они – племянники, разошедшиеся с людьми примерно 600 тысяч лет тому. В Европе они жили начиная с 250 тысяч лет назад, и исчезли – случайно или нет – после появления там людей, примерно 30 тысяч лет назад.
В последнее время чуть ли не потоком пошли научные статьи, противоречащие одна другой в отношении того, является ли неандерталец нашим предком. И несмотря на то, что был расшифрован геном неандертальца, точнее, неандерталки, вопрос так и остался нерешенным. Здесь просматривается занятная корреляция. Чем более пресса «желтая», тем больше она педалирует, что неандертальцы – наши предки. Интересно, почему? В общем, ответ ясен – выше продажи издания. В одной академической статье было найдено, что в нас содержится от 1% до 4% генома неандертальца. При этом авторы не указали, что погрешность этого определения не менее плюс-минум 10%. Но отметили, что полученные цифра очень предварительные, и нуждаются в перепроверке. Но прессу уже было не остановить. Да и в «академической» прессе часто встречаются ссылки на эти цифры, правда, про то, как получены и про погрешности – ни слова. Как и то, почему раньше они предками не были (см. диаграмму выше), а потом вдруг стали. В науке положено подвергать анализу все варианты «за» и «против», но так то в настоящей науке...
Да, обратно к «иммиграции» моих предков. Довольно легкое объяснение, больше журналистского характера, следующее. Первая волна в Европу (точнее, небольшие волны на протяжении 20 тысяч лет) оставила там шесть гаплогрупп. Носители первой, гаплогруппы U, дети воображаемой праматери «Урсулы», осели на территории будущей Греции 45 тысяч лет назад. Сейчас ее прямыми потомками являются примерно 11% современных европейцев, особенно в Англии и Скандинавии. Вторая, гаплогруппа X, праматерью которой была «Ксения» 25 тысяч лет назад, представлена шестью процентами европейцев, но ее ветви протянулись в Центральную Азию и Сибирь, и из Сибири – в Северную и Южную Америки. Около 1% американских индейцев – прямые потомки Ксении. В Европе живут три ветви прямых потомков «Ксении» - в Восточной Европе, в Центральной Европе, и в Англии.
Третья гаплогруппа H, или Helen, Елена, появилась в Европе 20 тысяч лет назад на границе современных Франции и Испании. Ее прямыми потомками являются 47% европейцев, включая мою маму, а значит, и меня. Генетики пока не знают, чем вызван такой успех потомков Елены по «завоеванию Европы» - то ли какими-то биологическими преимуществами, то ли просто так фишка легла. Будем считать, что второе.
Четвертая гаплогруппа V, прямые потомки «Велды», осели в Северной Испании 17 тысяч лет назад, и затем двинулись на север по следам уходящего ледника. 5% европейцев – прямые потомки «Велды», включая этническую группу Саами в Финляндии и Северной Норвегии.
В то же время, 17 тысяч лет назад, на Средиземноморье, в Северной Италии обосновалась пятая гаплогруппа Т, праматери «Тары». Сегодня примерно 9% европейцев – прямые потомки «Тары», они расселились вдоль Средиземного моря, и продвинулись в Англию и Ирландию.
Шестая гаплогруппа К с праматерью «Катрин» прибыли в Европу 15 тысяч лет назад и обосновались на южных склонах итальянских Альп. Недавно найденный в Альпийских горах «снежный человек» Отци, погибший 5 тысяч лет назад – прямой потомок «Катрин». Примерно 6% европейцев сегодня – прямые потомки «Катрин», в основном живут в Средиземноморье.
Наконец, седьмая, она же последняя гаплогруппа J с праматерью «Жасмин», пришедшие в Европу «второй волной» с территории нынешней Сирии уже после завершения великого ледникового периода, составляет примерно 17% современной Европы, включая Испанию и Португалию, Уэльс и Шотландию, и Центральную Европу. Эта волна прибыла в Европу относительно недавно, несколько тысяч лет тому, когда жители Ближнего Востока, озабоченные жесткой конкуренцией за земельные угодья, двинулись развивать сельское хозяйство в Европе, жители которой занимались в основном охотой. Они, эти новые переселенцы, слегка запутали генеалогическую картину Европы, но поскольку их в итоге стало несколько меньше одной пятой от уже порядком размножившихся коренных европейцев (точнее, европеек, говоря о мтДНК), то запутали не очень сильно.
ПОСЛЕСЛОВИЕ
Тема, безусловно, интересна. И понятно, почему – наша история, и тем более связь нашей личной истории с историей человечества не может не интересовать человека разумного – Нomo sapiens. Вспомните, когда вы впервые практически познакомились с системой компьютерного поиска, что вы немедленно выстучали на клавиатуре? Правильно, вашу фамилию.
Когда компания FamilyTreeDNA несколько лет назад прислала список моих маркеров Y-хромосомы и мтДНК и соответствующие гаплотипы, а также индексы гаплогрупп по мужской и материнской линии, для меня это была почти китайская грамота. И это несмотря на то, что мое образование не так далеко от этого. Что уж говорить о других! Дело в том, что совместное развитие генетики и генеалогии происходит так стремительно, что публикации и уровень знаний в этой области даже за 2000-й год представляют собой «каменный век» этой науки. Число обнаруженных маркеров в Y-хромосоме за последние два года выросло с 53 до многих сотен. Как я уже писал выше, последние два года число мтДНК в основной базе данных прибавляется по пять в день.
Еще совсем недавно было достаточно знать свой гаплотип (число повторов каждого маркера) всего для шести маркеров, чтобы считать эту информацию вполне исчерпывающей. Когда мне предложили сделать анализ на выбор по 12, 25 или 37 маркерам Y-хромосомы, я выбрал средний вариант, и, видимо, не ошибся. Потому что потом нашел полное совпадение своего гаплотипа с несколькими людьми в базе данных по 12 маркерам, но ни одного полного – по 25 маркерам (на самом деле был один, но с двумя генеалогическими шагами в сторону). Иначе говоря, 12 маркеров было еще недостаточно, но 25 маркеров – в самый раз. 37 маркеров ничего бы нового тогда не дали. Потом у меня определен 67-маркерный, а затем и 111-маркерный гаплотип, хотя самый ближний ко мне «родственник» так и остался на уровне 25-маркерного гаплотипа с двумя мутациями.
Но с тех пор, за последние два года я осознал, что самое увлекательное дело ДНК-генеалогии – это далеко не только поиски родственников. На самом деле 25-, 37-, 67-, и 111-маркерные гаплотипы позволяют построить «дерево гаплотипов» для широкого круга людей одного с нами рода, в моем случае – рода R1a, праславянского рода, и изучать историю рода по гаплотипам. Об этом – подробные рассказы в этой книге.
Для меня представило большое удовольствие распутать клубок, предоставленный мне ДНК-генеалогической компанией, и проследить исторический путь моих предков из Европы в Южную Сибирь и обратно в Европу. Если честно, то распутывая этот клубок, я переживал за них, они вдруг увиделись мне совершенно реальными, живыми людьми, со своими судьбами. Совершенно невероятно, как они, передвигаясь небольшими группами, сумели сохранить и пронести через десятки тысяч лет свой (наш!) семейный гаплотип, который записан в каждой из моих клеток. Работая над этим материалом, я вдруг ощутил причастность к нашей многотысячелетней семейной истории, причастность совершенно другого порядка, чем ощущал до сих пор.
Я совершенно ясно увидел, как моя древняя семья, мой семейный клан в количестве всего нескольких десятков человек – а тогда людей во всем мире было не более нескольких тысяч (не считая тех, кто уже давно жили в Африке, не будучи нашими предками) – отчаянно спорили, оставаться ли им на Равнине, или уходить навстречу солнцу. Они с болью разделились – одни остались, а другие ушли. И выжили в итоге те, кто ушли. Потом был другой переломный момент, в Алтайском регионе, когда клан опять разделился. И те, кто ушли, достигли – в своих потомках – Европы, а затем и территории будущей России. Те, кто остались – стали монголоидами, что само по себе и неплохо, но почти все погибли примерно 850 лет назад, и сейчас там – уже новые, монголоидные поколения R1a. Прошли бутылочное горлышко популяции, и сколько таких горлышек было у наших предков... Мы – потомки тех, кто чудом выжил бесчисленное число раз. Можно только представить, как много семей не выжили, и следов их сейчас нет. Нет, наверное, представить все-таки невозможно.
История воистину записана в наших ДНК. И то, о чем я здесь рассказываю, дает возможность ее ощутить по-новому, в буквальном смысле.
Желанием поделиться, желанием пригласить других испытать те же чувства и продиктован этот рассказ.
Должен сказать, что историю заселения Европы потомками семи женщин, праматерей основных европейских гаплогрупп, я позаимствовал из книги Брайана Сайкса «Семь Дочерей Евы» (W.W.Norton & Company, New York, London, 2001), хотя пересказов этой истории имеется предостаточно в генетической литературе. Все остальное в таком виде читатель нигде не найдет. Я бы хотел найти, и это сэкономило бы мне массу времени при интерпретации моих гаплотипов, но найти не смог. Более того, в ходе поисков понял, что и не найду. Потому и написал.
Анализ моей ДНК делала американская компания Family Tree DNA, http://www.familytreedna.com/, цены можно узнать здесь - http://www.familytre.../products.aspx. Подобные анализ делают многие компании в Европе, но я бы рекомендовал Family Tree. Они по запросу направляют вам маленькую щеточку типа зубной, ей надо поскрести примерно минуту за щекой, вложить в пластиковый футлярчик, вложить в прилагаемый конверт и бросить в почтовый ящик. Компания вам ответит.
А потом – интересное путешествие во времени.
Успеха.







0 Комментарии