Рис. 5
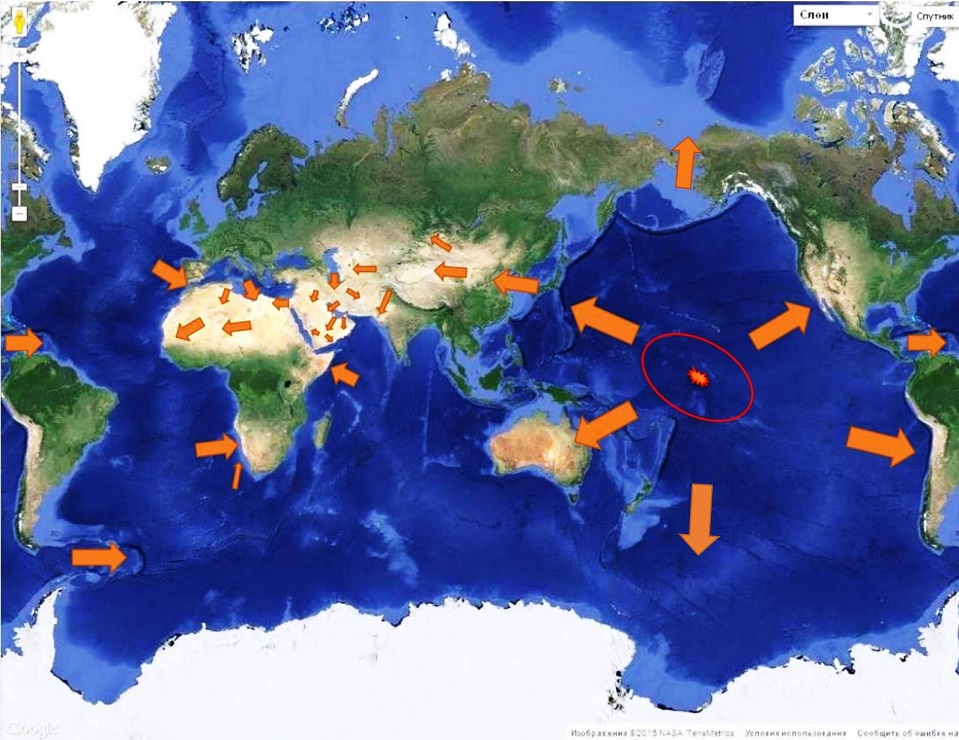
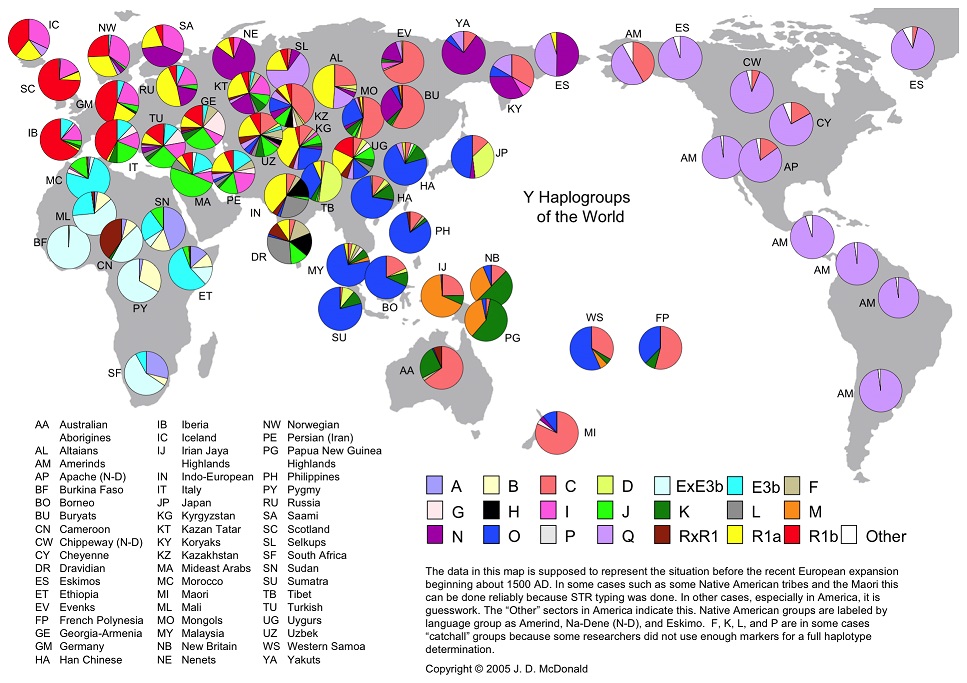
Гаплогруппа E, судя по широчайшему её распространении ныне в Африке (рис. 5), после отделения двинулась на юг, неся с собой свой вариант того же языка. Его реликт, подобный тому, чем на Дальнем востоке является язык айнов, также сохранился в Африке, как и гаплогруппа E. К нему относится язык «загадочных» гарамантов, о которых идёт речь в статье А. Милитарева «Глазами лингвиста: Гарамантида в контексте североафриканской истории», являющейся послесловием к книге «ГАРАМАНТИДА (африканская Атлантида)» (Рощин, 1994). Их современными наследниками являются туареги, сохранившие первозданную европеоидную антропологию и язык, сохранивший, как и язык айнов, славянскую корневую основу. Они до сих пор используют доныне звучащее письмо тифинаг, представляющее собой кальку славянской руницы. Их современный флаг доныне несёт на себе славянскую руну «Правда» - одну из, так называемых, рун-сутей славян. Материала шквал, и, если будет желание, огромную фактологию по гарамантам-туарегам, входящих в семью берберов Африки, можно обсудить в рамках Славянской академии. Исследованию праславянско-гарамантских соответствий посвящена статья «Мы вспомним всё», опубликованная в издании «Труды Профессорского клуба ЮНЕСКО»; Выпуск 8-9; Владивосток; 2004 г. (Юрковец, 2004).
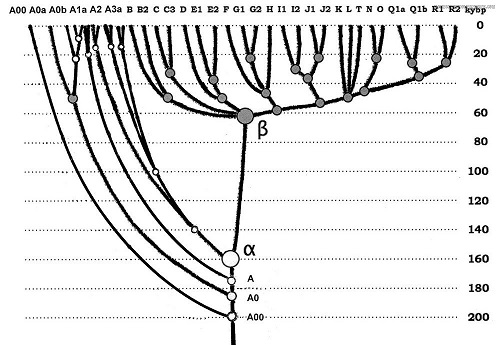
На представленной выше иллюстрации возраст DE представлен с большой ошибкой, поскольку выполнен в 2008 году (Википедия), когда надёжных способов определения возраста ветвей ещё не было. Позднее ошибка была исправлено в совместной работе А. Клёсова и И. Рожанского в 2013 году, как сказано выше.
Послесловие. Итак, согласно историческим, палеоклиматическим, ДНК-генеалогическим и лингвистическим данным, начало возрождения современного человечества уходит во времена, наступившие после космогенного катаклизма, имевшего место 64 +/- 6-7 тысяч лет назад и стёршего с лица земли предшествующую цивилизацию. Как показывают данные ДНК-генеалогии, в этом катаклизме смогла выжить единственная ветвь человечества, представленная гаплогруппой «бета» (В-Т). Её филогения укоренена на Среднедунайской низменности в пределах этой же даты и являющаяся единственной территорией на нашей планете, не затронутой космогенным мегацунами.
Как показали самые древние реликты индоевропейских языков (язык айнов и гарамантов) на этой территории жили будущие славяне (протославяне). Где они и живут в своём развитии все эти десятки тысяч лет вплоть до настоящего времени, всё это время оставаясь единой ветвью протославян-праславян-славян. Исходя из этого необходимо констатировать, что начиная с этой даты (64+/- 6-7 тысяч лет назад) человечество по мужской линии является единым. Т.е., глобализация по факту состоялась многие десятки тысяч лет назад и все мы давным-давно друг другу родственники по мужской линии, несмотря на столь, казалось бы, существенные различия между нами в виде видовых, расовых и национальных признаках.
Что касается собственно видового, расового и всех видов национального многообразия современного человечества – это, как оказалось, вклад женщин в современную антропологию. Этот вклад маркирует митохондриальная ДНК (мтДНК). Начало этой темы положено в работе «Кратко о палеоистории» в Вестнике Академии ДНК-Генеалогии, Т. 13; № 2; 2020 г. (Юрковец, 2020). При этом филогения женских мтДНК гаплогрупп, в отличие от мужской Y-ДНК, не показывает столь радикального перекоса численности в пользу какой-либо из ветвей, они более-менее равномерно покрывают собой всю планету, в крайнем случае подразделяясь на основные, малые и минорные. Доминирующей мт-гаплогруппы в мире нет, рис. 6.
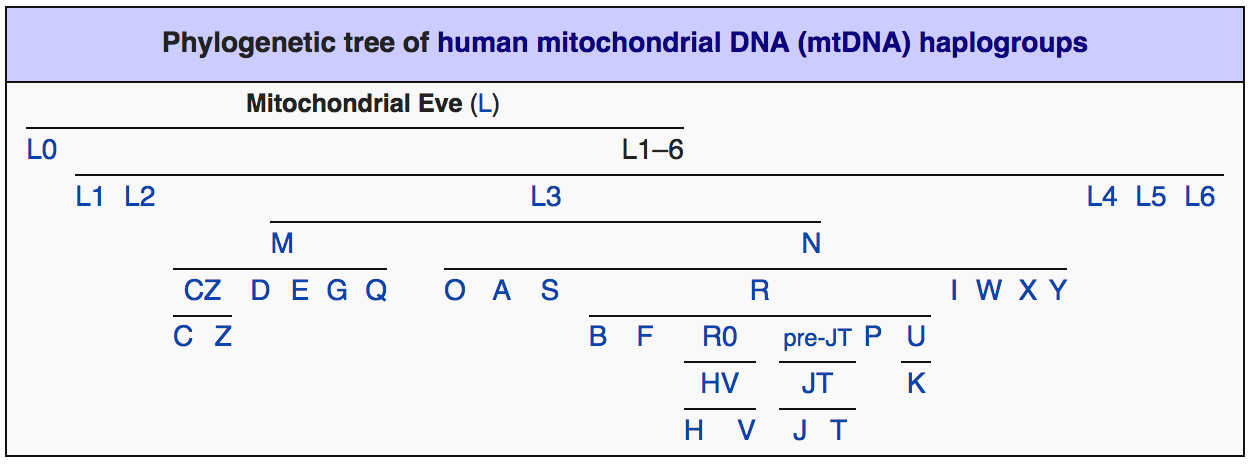
Рис. 6.
Принципиальное же отличие распределения женских гаплогрупп по планете от мужских состоит в том, что оно показывает полную картину расового разнообразия человека. Полную настолько, что его детальность, согласно филогенетическому древу женских мтДНК гаплогрупп превосходит любую из существующих расовых классификаций, рис. 6,. Такой «максималистский» вывод следует из довольно жёсткой территориальной привязки расовых ареалов к распространению определённых мтДНК гаплогрупп. Так, мт-гаплогруппа D численно преобладает на территории, вмещающей ядро монголоидной расы – в Восточной Азии. Распространение австралоидной расы, в основном, контролирует мт-гаплогруппа N в Австралии. Гаплогруппа M коррелирует с южноазиатской расой, территориально перекрываясь с мт-гаплогруппой D. Негрскую расу вмещает территория распространения субкладов негрской мт-гаплогруппы L в Африке. В Евразии и северном Средиземноморье - в пределах распространения европеоидной расы, доминируют две конкурирующие между собой по численности европеоидных мт-гаплогруппы – H и U. Мт-гаплогруппа H преобладает на западе, мт-гаплогруппа U – на востоке их общего евразийского ареала. Что, к слову, почти повторяет распределение мт-гаплогрупп M и D в Южной, Восточной и Юго-Восточной Азии.
С малыми расами не менее очевидную корреляцию обнаруживают столь же «малые» по численности гаплогруппы. Например, в Южной Африке южноафриканскую (бушменскую, капоидную, койсаноидную) малую расу контролирует мт-гаплогруппа L1. В Таиланде и Индии значительную долю составляет мт-гаплогруппа R, представляющая, по всей видимости, веддоидную малую расу. В Восточной и Юго-Восточной Азии – в Монголии, Китае, Японии, на Тайване, в Новой Гвинее и Полинезии заметную долю (до трети и более) составляет гаплогруппа B, коррелирующая с полинезийской малой расой.
В Евразии и южном Средиземноморье, в пределах распространения европеоидной расы, относительно равномерно распределена «малая» мт-гаплогруппа T, присутствующая, однако, здесь в заметных количествах. Здесь же, в Европе выделяются ещё четыре (кроме H, U и T, перечисленных выше) «дочери Евы» - J, K, X, V (Клёсов, 2016), которые, очевидно, будут «участвовать» в расовом разнообразии европейцев, когда будет выполнен поиск расовых соответствий с ними.
Существует ещё целый ряд минорных ветвей, рис. 6, которым, надо полагать, по итогам дальнейших исследований найдутся свои «минорные» расовые (?) соответствия. Конечно, за десятки тысяч лет изначальные расово-митохондриальные ареалы в значительной мере перемешались, их границы подверглись существенному размыванию, однако общая картина первоначального распространения рас и изначально несущих их мт-гаплогрупп всё ещё остаётся читаемой, а границы более-менее реконструируемыми, рис. 4.
Хронология мт-филогенеза, согласно существующему консенсусу (Soares, et al, 2009), берёт начало около 200 тысяч лет назад. Т.е., формирование рас на «подконтрольных» перечисленным выше мт-гаплогруппам территориях уходит в прошлое во времена архантропа (Дробышевский, antropogenez.ru), отличавшегося, судя по перечисленным выше корреляциям, таким же расовым разнообразием, как и современный человек и являющегося, таким образом, его – расового разнообразия – источником. Это подтверждают и археологические данные (Вейденрейх, Бунак, и др). Так, «прародитель» монголоидной расы архантроп синантроп, обнаружен в Восточной Азии – на территории распространения мт-гаплогруппы D. Питекантроп (также таксон «архантроп», как и все перечисленные далее, кроме палеоантропа неандерталенсиса), «источник» южноазиатской расы, найден на территории распространения гаплогруппы M – в Юго-Восточной Азии. Родезийский человек, «отец-основатель» большой негрской расы, когда-то существовал на территории нынешнего обитания мт-гаплогрупп L. Гейдельбергенсис (европейский эректус) - «патриарх» европеоидной расы, обитал на территории мт-гаплогруппы H, доминирование которой сейчас наиболее отчётливо проявлено в Западной Европе. Всё известное ныне разнообразие неандертальца выявлено в пределах географического ареала мт-гаплогруппы U, наследующей, по-видимому, его расовые особенности.
Таксон неандертальца – палеоантроп - казалось бы, несколько усложняет нарисованную выше картину тем, что обладает рангом, эволюционно превышающим ранг архантропа, от которого наследует расы современный человек. Однако нижняя граница хронологического интервала существования неандертальца как вида упирается в ту же датировку - 200 тысяч лет назад (Вишняцкий, 2010), где «легитимность» его таксона заканчивается. Из чего, как минимум, следует, что в части таксономии рода Homo не всё ещё определено и строго обосновано. Кроме того, антропология, как наука не только о человеке, но и о влиянии среды обитания на его эволюцию, в своих построениях не учитывает катастрофу, случившуюся 64 +/- 6-7 тысяч лет назад, которая какую-то часть ветвей тогдашнего человечества уничтожила, а остальных провела через бутылочное горлышко. А всё, что могло остаться от прекративших своё существование, скрыло под мощными наслоениями дилювия, ныне слагающего специфические (несущие диагностические признаки прохождения мощного потока) формы геологических тел (Юрковец, 2015). Поэтому пока всё, что мы сейчас можем констатировать, глядя на рис. 4, это то, что больше 130 тысяч лет нашей, пунктирно просматривающейся на 200 тысяч лет в прошлое истории, нам большей частью неведомо.
Литература:
Юрковец В.П. 2010. Климатические корреляции. Proceedings of the Academy of DNA Genealogy. Raleigh, N.C., Lulu. Vol. 3, Vol. No. 2. С. 301 – 325. ISSN 1942-7484
Юрковец В.П. 2012. Климатические катастрофы и история миграций основных гаплогрупп мужской половины человечества. Proceedings of the Academy of DNA Genealogy. Raleigh, N.C., Lulu. Vol. 5, Vol. No. 5. С. 568 – 586. ISSN 1942-7484
Клёсов А.А., Рожанский И.Л. 2011. Архаичная (архантропная в терминах антропологии) линия гаплогруппы А. Proceedings of the Academy of DNA Genealogy. Raleigh, N.C., Lulu. Vol. 4, Vol. No. 7. С. 1367 – 1494. ISSN 1942-7484
Bassinot F., Labeyrie L., Vincent E., et al. 1994. The astronomical theory of climate and the age of the Brunhes-Matuyama magnetic reversal // Earth Planet. Sci. Lett. V. 126, P. 91-108
Козелков А.С., Куркин А.А., Пелиновский Е.Н. 2014. Цунами космогенного происхождения // Труды Нижегородского государственного технического университета им. Р.Е. Алексеева № 2(104). С. 26-35.
Юрковец В.П. 2015. Климатическая катастрофа гаплогруппы «бета» //
Proceedings of the Academy of DNA Genealogy. Raleigh, N.C., Lulu. Vol. 8, No. 3. -P. 376-432.ISSN 1942-7484.
Костриков А.А. Геофизическая геодинамика сверхмощных потоков ледникового периода // Материалы гляциологических иследований, 2003. Вып. 95. С. 22–27.
Юрковец В.П. 2013. Кратко о палеоистории // Proceedings of the Academy of DNA Genealogy. Raleigh, N.C., Lulu. Vol. 13, No. 2. C. 215-222. ISSN 1942-7484
Юрковец В.П., Василенко С.И. 2011 ДНК-генеалогия, палеоклимат и геоморфология. Гидродинамический карст. 2010 Proceedings of the Academy of DNA Genealogy. Raleigh, N.C., Lulu. Vol. 10, Vol. No. 3. С. 1412 – 1442. ISSN 1942-7484
Клёсов А.А. 2011. ДНК-генеалогия основных гаплогрупп мужской половины человечества (Часть 2). Proceedings of the Academy of DNA Genealogy. Raleigh, N.C., Lulu. Vol. 4, Vol. No. 7. С. 1367 – 1494. ISSN 1942-7484
Клёсов А.А. 2016. ДНК-генеалогия от А до Т. Лаборатория ДНК-Генеалогии. ISBN 978-5-8041-0873-2 – 475
Вишняцкий Л.Б. Неандертальцы: история несостоявшегося человечества. / — СПб. : Нестор-История, 2010. — 312 с. ; ил.
Дробышевский С.В. Судьба Неандертальцев. Сайт Антропогенез.ру. Режим доступа: https://antropogenez.ru/article/239/
Таксами Ч.М., Косарев В.Д. Кто вы, айны? – М.: Мысль, 1990.- 318, [2] c. ISBN 5-244-00431-X
Юрковец В.П. Славянские руны. – СПб.: Родострой. 2004. № 4. С. 14 - 16
Клёсов Анатолий. Народы России ДНК-Генеалогия. - СПб.: Питер, 2021. – 784с.: ил. ISBN 978-5-00116-595-8
Павленко Н.И. Штрихи к познанию языка айнов. 2000. ВЕСТНИК САХАЛИНСКОГО МУЗЕЯ. № 7. С. 331-333. Вестник сахалинского областного краеведческого музея. Южно-Сахалинск