
В Вестнике Академии ДНК-генеалогии, т.13, № 2, вышла статья "Кратко о палеоистории", которая является началом этой - о палеоистории - темы. Первая часть посвящена проблеме происхождения рас, которая, решается на стыке нескольких дисциплин: палеогеографии, ДНК-генеалогии, антропологии и археологии.
Статья написана при непосредственном участии Анатолия Алексеевича Клёсова. В ней есть вставки его "прямой речи" из наших дискуссий, которые мы вели по почте на каждом повороте темы. Фактически Анатолий Алексеевич является соавтором, но без его согласия я не могу поставить его фамилию в оглавлении статьи. Быть может, наша дискуссия ещё не закончилась?
КРАТКО О ПАЛЕОИСТОРИИ
Обнаружение не так давно следов прохождения космогенного потока на сооружениях Древнего царства Египта открыло тему палеоистории (Юрковец, 2015). Т.е. истории существования некоей древней цивилизации, существовавшей на нашей планете до планетарного катаклизма, проведшего 67+/-5 тысяч лет назад через жёсткое «бутылочное горлышко» предшествующее человечество (Клёсов, 2011). В настоящее время в профильных публикациях в сети появляется всё больше материалов, из которых следует, что т.н. «гибель мегафауны» в позднем плейстоцене также была неизбежным следствием этого космогенного катаклизма, и заключалась она, видимо, не только в прямом уничтожении гигантов, но и в разрушении привычной среды обитания, разрывах пищевых цепочек, глобальном похолодании, наступившем в результате выброса в атмосферу огромного количества продуктов импакта, а также вулканического пепла из разбуженных ударом из космоса недр (Юрковец, Василенко, 2017). В антропологии пока не принято рассматривать человека, как свидетеля и участника этой вселенской драмы, несмотря на то что она никак не могла обойти его стороной. В настоящей работе будут приведены новые данные, из которых постепенно начинает складываться мозаика во многом кажущихся фантастическими деталей предшествующего мира, случившейся катастрофы и её, пока не вмещающихся в современные представления о мире, последствий. Этих данных с каждым годом становится всё больше, однако нам, несмотря на наши знания о планетарных катаклизмах в геологическом прошлом Земли, в том числе, не столь уж далёком - сопоставимом с временем существования человека, по-прежнему кажется, что «это» было где-то «там», а не с нами. Однако, выводы, например, той же ДНК-генеалогии, касающиеся последних 200 тысяч лет биологической истории нашего вида, показывают, что это далеко не так. С них и начнём.
По данным ДНК-генеалогии, подавляющая часть всех современных мужчин мира - почти 100%, независимо от рас и мест обитания, включая Африку, относится к потомкам гаплогруппы «бета» (гаплогруппа B-T), прошедшей «бутылочное горлышко» 67 +/-5 тысяч лет назад (Клёсов, 2011). Понятно, что когда-то существовавший общий предок этих, почти всех ныне существующих мужчин мира относился к одной расе, видимо, к европеоидной, просто исходя из того, что производные от него древнейшие ветви филогенетического древа Y-хромосомы человека, в основном, европеоидные (Е, G, I, J, R, T), а всё современное физическое разнообразие человечества за 64 тысячи лет «с нуля» образоваться не могло, слишком большие различия в десятках антропологических показателей (Клёсов).
Предков гаплогруппы «бета» пока не найдено нигде, в том числе в Африке, где обнаружены потомки самой древней из всех известных мужских гаплогрупп мира - A00, проживающей ныне только здесь, на этом континенте (Клёсов, Рожанский, 2011). Из этого следует, что, во-первых, место прохождения гаплогруппой «бета» «бутылочного горлышка» не Африка. Филогенетическое древо, показанное на рис. 1, это запрещает. Во-вторых, то, что древнейшая мужская гаплогруппа мира A00 из Африки никогда не выходила. Однако в глубокой древности зайти в Африку она могла. Филогенетическое древо это позволяет, хотя и не обязывает. Поэтому эти две неравных, как по численности, так и по возрасту, ветви условно названы «неафриканской» (первая) и «африканской» (вторая). Условно потому, что, строго говоря, неизвестно, где образовались они обе – Филогенетическое древо Y-хромосомы человека не укоренено. Забегая вперёд, можно сказать, что место прохождения гаплогруппой «бета» «бутылочного горлышка (территория выживания в космогенном катаклизме) известно, об этом далее.
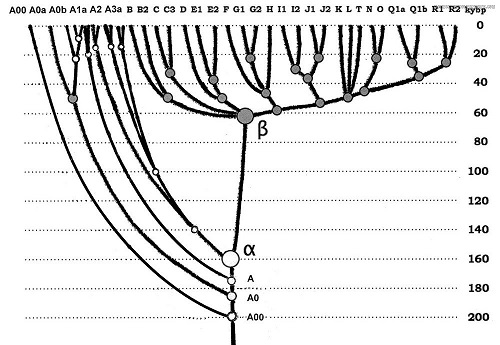
Рис. 1. Филогенетическое древо Y-хромосомы (Клёсов, Рожанский, 2013)
Общий счёт набежавших мутаций между «африканскими» и «неафриканскими» ветвями уводит их общего предка более чем на 200 тысяч лет назад, рис. 1. При этом, сами «африканские» ветви представлены короткими обрывками малой численности, общие предки которых уходят в прошлое не далее тысячи с небольшим лет (Клёсов, 2016). Поэтому пока всё, что можно извлечь из их гаплотипов, относится исключительно к эволюционным отрезкам времени и ничего не говорит об истории и географии их миграций.
Возраст прохождения «бутылочного горлышка» гаплогруппой «бета» (B-T) совпадает с датой падения массивного космического тела в Тихий океан, определённой по сумме палеогеографических, палеобиологических и палеоклиматических данных (Юрковец, 2015). По этим же данным уверенно определяется место прохождения ей «бутылочного горлышка», которое находилось на территории Среднедунайской низменности, со всех сторон закрытой высокогорными системами Центральной Европы: Западными, Восточными и Южными Карпатами, Стара-Планиной, Динарским нагорьем и Восточными Альпами (Юрковец, Василенко, 2017). Т.о. причиной прохождения «бутылочного горлышка» является космогенный катаклизм, а показанное ниже на рис. 2 современное мировое распределение Y-гаплогрупп является итогом почти 70-тысячелетних миграций этой практически единственной выжившей в космогенном катаклизме ветви.
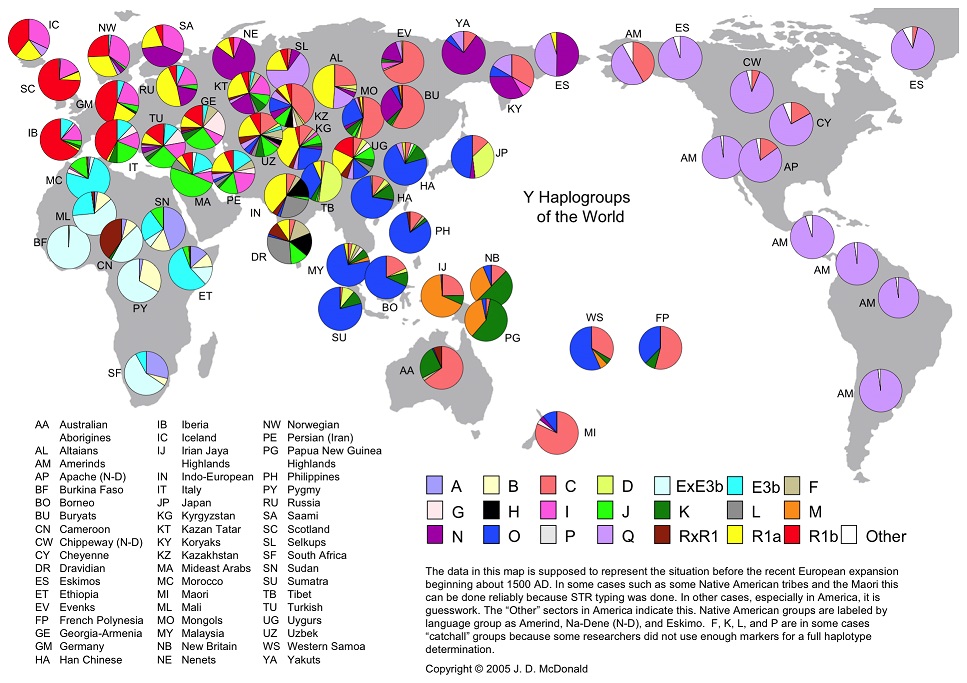
Рис. 2. Y-гаплогруппы мира (из открытого доступа в интернете)
Среднедунайская низменность располагается на обратной от места падения космического тела в глубоководную часть Тихого океана стороне земного шара, поэтому гигантская волна пришла сюда максимально ослабленной и не смогла преодолеть здешние вершины. В итоге на этой территории выжили не только европейские представители рода человеческого, но и вся обитавшая здесь флора и фауна, включая т.н. «мегафауну», обеспечив, таким образом, материальный базис для выживания и последующего возрождения человечества.
Филогения женских мт-ДНК гаплогрупп, наоборот, не показывает столь радикального перекоса численности в пользу какой-либо из ветвей, они более-менее равномерно покрывают собой всю планету, в крайнем случае подразделяясь на основные, малые и минорные. Доминирующей мт-гаплогруппы в мире нет, рис. 3.
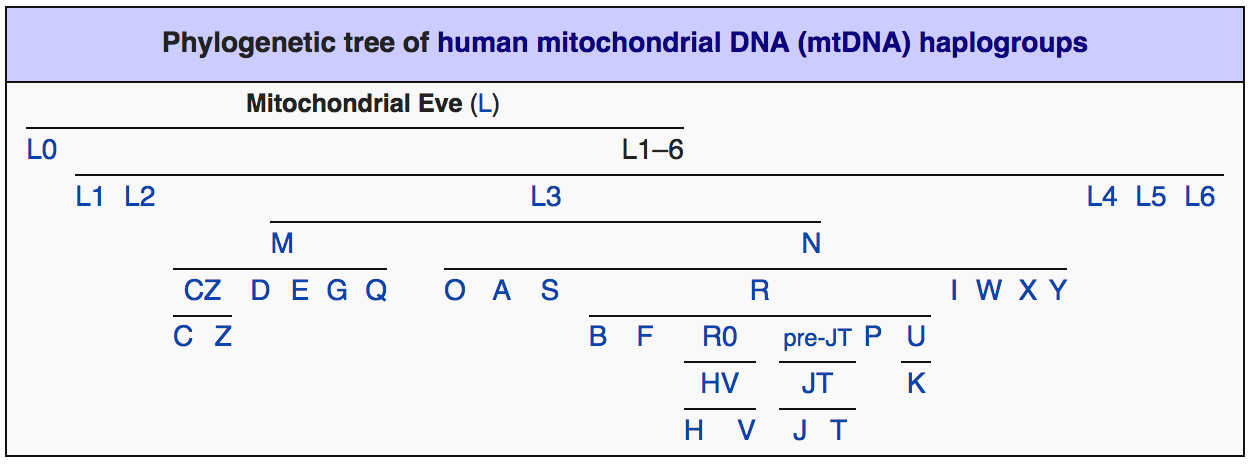
Рис.3. Филогенетическое древо мт-ДНК гаплогрупп (из открытого доступа в интернете)
Географическое распространение женских мт-ДНК гаплогрупп показано на рис. 4.
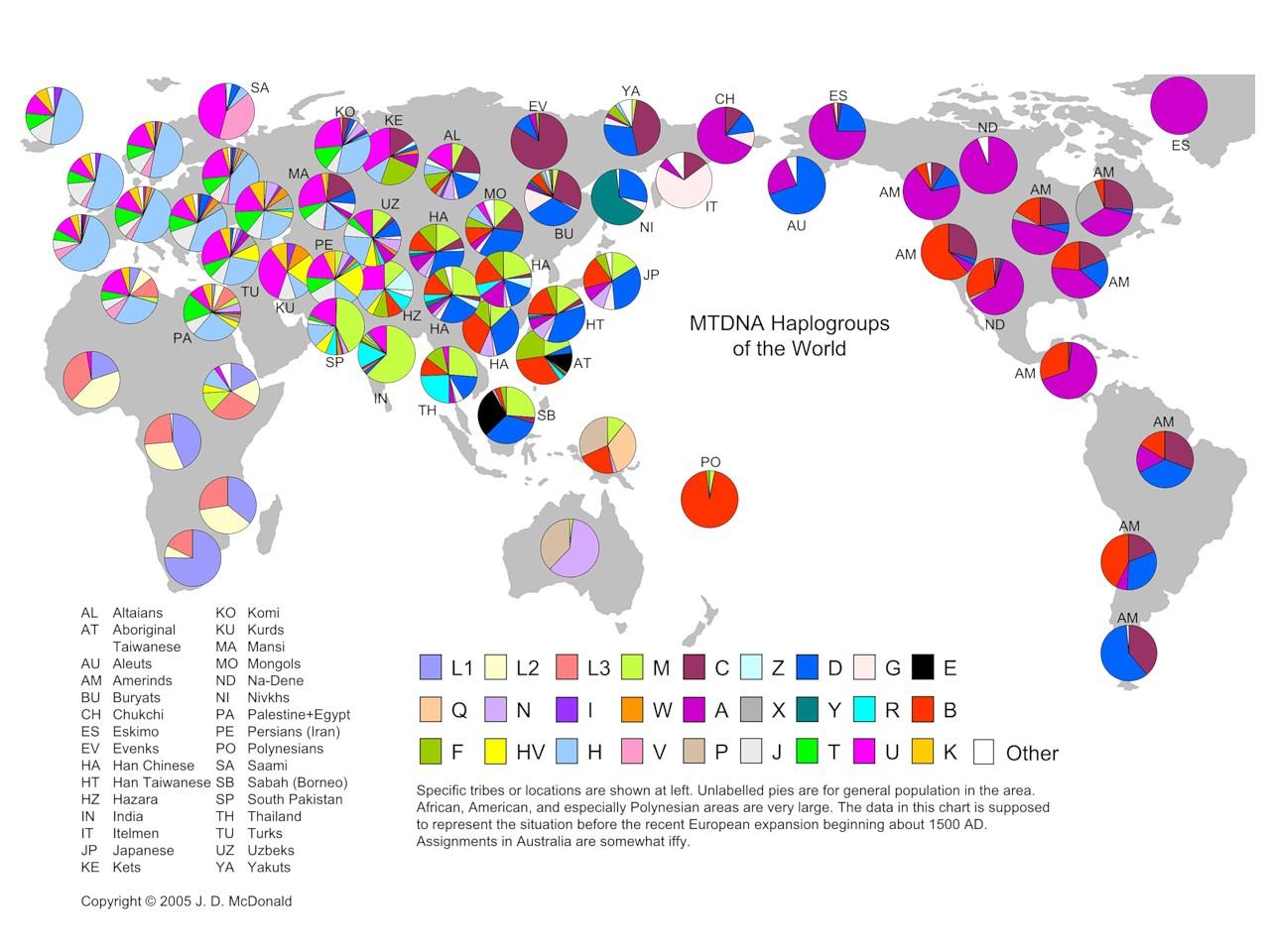
Рис 4. Мт-ДНК гаплогруппы мира (из открытого доступа в интернете)
Первое, что обращает на себя внимание в распространении как мужских, так и женских ветвей, это минимальную вариабельность и тех, и других по периферии обитаемой ойкумены, центром которой со всей очевидностью – не только по максимуму разнообразия, но и чисто географически, выступает зона умеренного климата Евразии. Что вполне объяснимо – в этой зоне в течение всего антропогена наблюдались и наблюдаются максимальные амплитуды и количество климатических перемен - от оледенений и межледниковий, увлажнений и аридизаций до чередований времён года, наиболее контрастных именно здесь. Природа и климат постоянно гоняли и гоняют человечество в этой полосе с места на место, заставляя каждый раз изобретать всё новые способы выживания, стимулируя тем самым его развитие (Юрковец, 2011).
Принципиальное же отличие распределения женских гаплогрупп по планете от мужских состоит в том, что оно показывает полную картину расового разнообразия человека. Полную настолько, что его детальность, согласно филогенетическому древу мт-ДНК гаплогрупп, рис. 3, превосходит любую из существующих расовых классификаций. Такой «максималистский» вывод следует из довольно жёсткой территориальной привязки расовых ареалов к распространению определённых мт-ДНК гаплогрупп. Так, мт-гаплогруппа D численно преобладает на территории, вмещающей ядро монголоидной расы – в Восточной Азии. Распространение австралоидной расы, в основном, контролирует мт-гаплогруппа N в Австралии. Гаплогруппа M коррелирует с южноазиатской расой, территориально перекрываясь с мт-гаплогруппой D. Негрскую расу вмещает территория распространения субкладов мт-гаплогруппы L в Африке. В Евразии и северном Средиземноморье - в пределах распространения европеоидной расы, доминируют две конкурирующие между собой по численности европеоидных мт-гаплогруппы – H и U. Мт-гаплогруппа H преобладает на западе, мт-гаплогруппа U – на востоке их общего евразийского ареала. Что, к слову, почти повторяет распределение мт-гаплогрупп M и D в Южной, Восточной и Юго-Восточной Азии.
С малыми расами не менее очевидную корреляцию обнаруживают столь же «малые» по численности гаплогруппы. Например, в Южной Африке южноафриканскую (бушменскую, капоидную, койсаноидную) малую расу контролирует мт-гаплогруппа L1. В Таиланде и Индии значительную долю составляет мт-гаплогруппа R, представляющая, по всей видимости, веддоидную малую расу. В Восточной и Юго-Восточной Азии – в Монголии, Китае, Японии, на Тайване, в Новой Гвинее и Полинезии заметную долю (до трети и более) составляет гаплогруппа B, коррелирующая с полинезийской малой расой.
В Евразии и южном Средиземноморье, в пределах распространения европеоидной расы, относительно равномерно распределена «малая» мт-гаплогруппа T, присутствующая, однако, здесь в заметных количествах. Здесь же, в Европе выделяются ещё четыре (кроме H, U и T, перечисленных выше) «дочери Евы» - J, K, X, V (Клёсов, 2016), которые, очевидно, будут «участвовать» в расовом разнообразии европейцев, когда будет выполнен поиск расовых соответствий с ними.
Существует ещё целый ряд минорных ветвей, рис. 3, которым, вероятно, по итогам дальнейших исследований найдутся свои «минорные» расовые (?) соответствия. Конечно, за десятки тысяч лет изначальные расово-митохондриальные ареалы в значительной мере перемешались, их границы подверглись существенному размыванию, однако общая картина первоначального распространения рас и изначально несущих их мт-гаплогрупп всё ещё остаётся читаемой, а границы более-менее реконструируемыми, рис. 4.
Хронология мт-филогенеза, согласно существующему консенсусу (Soares, et al, 2009), берёт начало около 200 тысяч лет назад. Т.е., формирование рас на подконтрольных перечисленным выше мт-гаплогруппам территориях уходит в прошлое во времена архантропа (Дробышевский, antropogenez.ru), отличавшегося, судя по перечисленным выше корреляциям, таким же расовым разнообразием, как и современный человек и являющегося, таким образом, его – расового разнообразия – источником. Это подтверждают и археологические данные (Вейденрейх, Бунак, и др). Так, «прародитель» монголоидной расы архантроп синантроп, обнаружен в Восточной Азии – на территории распространения мт-гаплогруппы D. Питекантроп (также таксон «архантроп», как и все перечисленные далее, кроме палеоантропа неандерталенсиса), «источник» южноазиатской расы, найден на территории распространения гаплогруппы M – в Юго-Восточной Азии. Родезийский человек, «отец-основатель» большой негрской расы, когда-то существовал на территории нынешнего обитания мт-гаплогрупп L. Гейдельбергенсис (европейский эректус) - «патриарх» европеоидной расы, обитал на территории мт-гаплогруппы H, доминирование которой сейчас наиболее отчётливо проявлено в Западной Европе. Всё известное ныне разнообразие неандертальца выявлено в пределах географического ареала мт-гаплогруппы U, наследующей, по-видимому, его расовые особенности.
Таксон неандертальца – палеоантроп - казалось бы, несколько усложняет нарисованную выше картину тем, что обладает рангом, эволюционно превышающим ранг архантропа, от которого наследует расы современный человек. Однако нижняя граница хронологического интервала существования неандертальца как вида упирается в ту же датировку - 200 тысяч лет назад (Вишняцкий, 2010), где «легитимность» его таксона заканчивается. Из чего, как минимум, следует, что в части таксономии рода Homo не всё ещё определено и строго обосновано. Кроме того, антропология, как наука не только о человеке, но и о влиянии среды обитания на его эволюцию, в своих построениях не учитывает катастрофу, случившуюся 67 +/- 5 тысяч лет назад, которая какую-то часть ветвей тогдашнего человечества уничтожила, а остальных провела через бутылочное горлышко. А всё, что могло остаться от прекративших своё существование, скрыло под мощными наслоениями дилювия, ныне слагающего специфические (несущие диагностические признаки прохождения мощного потока) формы геологических тел (Юрковец, 2015). Поэтому пока всё, что мы сейчас можем констатировать, глядя на рис. 1, это то, что больше 130 тысяч лет нашей, пунктирно просматривающейся на 200 тысяч лет в прошлое истории, нам большей частью неведомо.
В том числе, и из-за нашего незнания, о чём будет отдельная глава не только о филогении мужских гаплогрупп «старого света», но и женских «нового», автохтоны которых там пока неизвестны, наравне с мужскими, что является существенным отличием в этой части «старого света» от «нового» и, похоже, свидетельствует об уничтожении катаклизмом большей части автохтонов «нового». Впрочем, положение с выживанием человека в космогенной катастрофе в «новом свете», судя по некоторой, не совсем достоверной информации, поступающей оттуда, не совсем безнадёжно, поэтому окончательные выводы ещё делать рано.
Разумеется, все расы на соответствующих им территориях обитания мт-гаплогрупп обязаны были когда-то иметь свои Y-хромосомные ветви, без которых они не смогли бы просуществовать ни одного поколения. Из этого следует неизбежный вывод – в космогенном катаклизме выжила не только европеоидная ветвь мужской половины человечества, а, как минимум, какая-то часть больших и малых её ветвей, бывших когда-то половинками ныне существующих митохондриальных гаплогрупп. Однако им по каким-то причинам (проигрыш в эволюционной «битве»?) не повезло и они, за исключением Y-гаплогрупп A00 – «альфа», не оставили потомства. Что, конечно, также не является «терминальным» выводом в силу тех же причин.
(продолжение следует)








